Способ создания форм сои с измененным жирно-кислотным составом маслаПатент на изобретение №: 2215406 Автор: Зеленцов С.В., Кочегура А.В., Шведов И.В., Шишков Г.З. Патентообладатель: Государстенное научное учреждение РАСХН - Всероссийский научно-исследовательский институт масличных культур им. В.С. Пустовойта Дата публикации: 10 Ноября, 2003 Начало действия патента: 3 Января, 2002 Адрес для переписки: 350038, г.Краснодар, ул. Филатова, 17, ГНУ РАСХН ВНИИМК, патентная группа Изображения Изобретение предназначено для использования в сельском хозяйстве. Способ включает перевод исходных диплоидных сортов на тетраплоидный уровень колхицинированием или другим известным способом, выделение реверсивных диплоидных растений по морфологическим и другим хозяйственно ценным признакам и использование их в качестве исходного материала. Полученные тетраплоиды выращивают в течение нескольких поколений и одновременно с этим в каждом поколении отбирают рекомбинированные тетраплоидные формы, отличающиеся от базового тетраплоидного морфотипа. Отобранные формы размножают каждую в отдельности в течение ряда поколений, в каждом из которых выделяют формы с реверсивно кратно-пониженной плоидностью (реплоиды), плоидность которых определяют по морфологическим признакам и прямым подсчетом хромосом. Оценивают реплоиды по жирно-кислотному составу масла, выделяя формы, отличающиеся от растений с исходной плоидностью измененным или новым жирно-кислотным составом масла. Изобретение позволяет создавать формы сои без использования мутагенов. 2 з. п.ф-лы, 1 табл. Изобретение относится к сельскому хозяйству и может быть использовано в селекции сои. Известен метод создания низколиноленового безэрукового ярового рапса, включающий межвидовую гибридизацию горчицы сарептской с безэруковым рапсом, индивидуальный отбор слабофертильных межвидовых гибридов F1, направленное переопыление с яровым безэруковым рапсом и выделение в F2 среди растений рапсового типа низколиноленовых форм (В. И. Шпота, В.Е.Подколзина. Метод создания низколиноленового безэрукового ярового рапса // Вестник с.-х. науки, 1982. - 7 (310). - С. 59-62). Недостатком известного способа является его узкая внутриродовая специфичность, позволяющая получать известные результаты только в пределах рода Brassica, отличающегося фертильностью гибридов при скрещивании видов с различным количеством хромосом. Согласно известному способу в межвидовую гибридизацию вовлекают культурные виды: горчицу сарептскую (Brassica juncea Czern. ) и яровой pane (Brassica napus oleifera Metzg.) с разным количеством хромосом (2n=36 у горчицы и 2n=38 у рапса). При этом гаплоидные гаметы в F1 будут иметь плоидность n= 18 или n=19, a гибриды F1 будут иметь горчичный (2n= 36), рапсовый (2n= 38) или промежуточный слабофертильный (2n=37) типы плоидности. Последующее переопыление (беккросс) слабофертильных межвидовых гибридов анеуплоидного типа (2n=37) с яровым рапсом (2n=38) приводит к созданию замещенных линий рапса (2n=38) с трансгрессивно измененным жирно-кислотным составом масла. Культурная соя (Glycine max (L.) Merr.) является единственным культурным видом в роде Glycine Willd. Успешная межвидовая гибридизация культурной сои (2n=40) возможна только с дикорастущей уссурийской соей (2n= 40, G. soja Sieb. and Zucc.), отличающейся от культурной сои комплексом хозяйственно ценных признаков, в т.ч. повышенным уровнем полиненасыщенных (линолевой и линоленовой) кислот, при этом межвидовые гибриды сои имеют такое же количество хромосом (2n=40), фертильны, а все морфологические и биохимические признаки (в т.ч. жирно-кислотный состав масла) у них наследуется промежуточно. В целом, межвидовая гибридизация у сои приводит к созданию полукультурных гибридов с несколько повышенным содержанием полиненасыщенных жирных кислот и не обеспечивает трансгрессивного изменения жирно-кислотного состава масла. Известно использование химического мутагенеза в селекции подсолнечника для изменения соотношения жирных кислот в масле, согласно которому обрабатывают семена 0,5%-ным раствором мутагена диметилсульфата, отбирают в М3 родоначальные высокоолеиновые растения с содержанием олеиновой кислоты около 50%, осуществляют индивидуальный отбор с оценкой по потомству и проводят переопыление лучших высокоолеиновых растений (К.И.Солдатов Использование химического мутагенеза в селекции подсолнечника / В сб.: Материалы VII международной конференции по подсолнечнику 27 июня - 3 июля 1976 г. / М.: "Колос". - 1976. - С. 179-182). Недостатком известного способа является то, что получение подсолнечника с измененным жирно-кислотным составом масла достигается методом химического мутагенеза, а использование физических или химических мутагенов в критических дозах (LD50) приводит к модификации многих хозяйственно ценных признаков за счет нарушения функциональности отдельных локусов, контролирующих их реализацию. При этом в геноме растения увеличивается доля морфологически незаметных мутаций и микромутаций, оказывающих аддитивно-негативное действие на жизнеспособность организма в целом и вызывая снижение продуктивности и масличности растений, а также устойчивости к патогенной микрофлоре. Их искусственная элиминация из генома требует многочисленных возвратных скрещиваний с другими формами и последующей оценкой потомства. Как правило, все признаки, полученные путем мутагенеза, в том числе и при использовании известного метода рецессивны и требуют постоянного поддержания организма в гомозиготном состоянии, что очень проблематично для видов с перекрестным типом опыления. Кроме того, многочисленные попытки в прошлом использовать известный метод для изменения жирно-кислотного состава масла у сои не привели к ожидаемым результатам. В целом, известный метод узкоспецифичен для подсолнечника и не вызывает существенных генетических модификаций жирно-кислотного состава масла у сои, полученные высокоолеиновые формы подсолнечника требуют пространственной изоляции от других форм и несут комплекс отрицательных мутаций, оказывающих негативное влияние на реализацию других хозяйственно ценных признаков. Известен также способ создания исходного материала для селекции зерновых и крупяных культур, выбранный нами в качестве прототипа. Известный способ включает перевод диплоидных сортов на тетраплоидный уровень колхицинированием или другим известным способом, обработку семян тетраплоидов 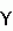 -излучением или раствором нитрозоэтилмочевины в дозе, критической для диплоидной формы, выращивание поколений M1 и M2 в виде популяций, выделение в поколении М2 реверсивных диплоидных растений (по следующим признакам: большая высота растений, раннее колошение, высокая озерненность колоса), использование семян с отобранных растений для закладки линий (а.с. СССР 1519596, А 01 Н 1/04, публ. 1989 г.). Хотя известный способ по названию не совпадает с названием заявляемого нами изобретения, он по сущности своей может быть использован для изменения жирно-кислотного состава масла, если всей совокупности приемов известного способа будет подвергнута масличная культура (например, соя), а среди полученных реверсивных диплоидных растений, оцененных по жирно-кислотному составу масла, будут выделены реверсанты с измененным соотношением жирных кислот. Недостатком известного способа, взятого нами в качестве прототипа, является то, что для создания форм растений с измененным соотношением жирных кислот в масле используются мутанты реверсивно диплоидного происхождения. Использование физических или химических мутагенов в критических дозах (LD50) приводит к модификации хозяйственно ценных признаков за счет нарушения функциональности отдельных локусов, контролирующих их реализацию. При этом в организме увеличивается количество негативных мутаций, оказывающих аддитивно-негативное действие на жизнеспособность организма в целом. Разнообразие реверсивных диплоидов при использовании -излучения или раствора нитрозоэтилмочевины определяется только морфологическими признаками (большая высота растений, раннее колошение, высокая озерненность колоса). Использующиеся в прототипе приемы для увеличения частоты появления реверсивных диплоидов существенно ограничивают возможности для отбора хозяйственно ценных диплоидных реверсантов, поскольку их отбор ведется только после обработки мутагенами в критических дозах (т.е. при летальности не менее 50% особей). При этом часть полезных генных рекомбинаций, в т.ч. и по жирно-кислотному составу масла, образовавшихся на полиплоидном уровне или при реверсии на диплоидный уровень, будет утеряна из-за образования несовместимых с жизнью организма мутаций, вызванных применением критических доз мутагенов, а большинство вновь образовавшихся признаков, в т.ч. и изначально доминантных у исходного сорта или тетраплоида, приобретут рецессивность из-за частичного повреждения и нарушения функциональности мутировавших локусов. В целом, известный способ, выбранный нами в качестве прототипа, существенно ограничивает получение, выявление и последующее сохранение максимально возможного полиморфизма по жирно-кислотному составу масла. Заявляемым изобретением решается задача повышения эффективности способа за счет расширения полиморфизма по соотношению компонентов масла при получении реверсантов (реплоидов) (впервые введенный нами термин) сои. Цель изобретения - создание форм сои с измененным жирно-кислотным составом масла без использования мутагенов различной природы, путем полиплоидизации растений, последующего отбора рекомбинантных полиплоидов и выделения среди них за счет спонтанной редукции плоидности гамет форм с возвратной кратно-пониженной плоидностью - реплоидов. Технический результат достигается тем, что в известном способе, включающем перевод исходных диплоидных сортов на тетраплоидный уровень колхицинированием или другим известным способом, выделение реверсивных диплоидных растений по морфологическим и другим хозяйственно ценным признакам и использование их в качестве исходного материала, согласно изобретению полученные тетраплоиды выращивают в течение нескольких поколений и одновременно с этим в каждом поколении отбирают рекомбинированные тетраплоидные формы, отличающиеся от базового тетраплоидного морфотипа, отобранные формы размножают каждую в отдельности в течение ряда поколений, в каждом из которых выделяют формы с реверсивно кратно-пониженной плоидностью (реплоиды), плоидность которых определяют по морфологическим признакам и прямым подсчетом хромосом, оценивают реплоиды по жирно-кислотному составу масла, выделяя формы, отличающиеся от растений с исходной плоидностью измененным или новым жирно-кислотным составом масла. Причем реплоиды сои выделяют по морфологическим, биохимическим или иным признакам, например выделяют реплоиды сои с повышенным или пониженным содержанием олеиновой, линолевой, линоленовой и других жирных кислот. Сопоставительный анализ заявляемого технического решения с прототипом позволяет сделать вывод, что заявляемый способ отличается от известного наличием нового приема: выделением рекомбинированных тетраплоидов из внутрисортовой популяции тетраплоидов, полученных удвоением плоидности исходных диплоидных сортов и выращиваемых в течение нескольких поколений, а также порядком выполнения известного приема "выделение реверсивных форм": формы с реверсивно кратно-пониженной плоидностью (реплоиды) выделяют из рекомбинированных тетраплоидных форм при их размножении в течение ряда поколений. Таким образом, заявляемый способ соответствует критерию патентоспособности "НОВИЗНА". Исследуя уровень техники в процессе проведения патентного поиска по всем видам сведений, общедоступных в печати, мы обнаружили, что способ создания форм сои с измененным жирно-кислотным составом масла, включающий заявленную нами совокупность признаков, для специалиста в области селекции явным образом не следует из известного на сегодня существующего уровня техники. Нашими исследованиями на протяжении ряда лет выявлено, что в природе с определенной частотой постоянно осуществляется спонтанная полиплоидизация высших растений. Полиплоидизация приводит к образованию поливалентов в профазе I мейоза I. С этим связаны различные нарушения конъюгации и кроссинговера гомологичных хромосом, вызывающие их структурные изменения. Нерегулярное распределение дочерних хромосом в мейозе у "ранних" автополиплоидов приводит к редукции плоидности гамет и образованию анеуплоидов и возвратных диплоидов (реплоидов). У "ранних" автополиплоидов при мультивалентной конъюгации и кроссинговере могут происходить асимметричные обмены гомологичными участками хромосом, сопровождающиеся дупликациями и делециями. Структурные изменения хромосом могут приводить к количественным и качественным изменениям существующих или к образованию новых признаков. Реплоидные формы образуются на основе составляющих полиплоидный геном, диплоидных геномов, включая рекомбинированные наборы хромосом. При этом образовавшиеся реплоиды могут быть гомологичными исходной диплоидной форме или отличаться от нее, сохраняя возникшие на полиплоидном уровне новые признаки или приобретая их при мультивалентной конъюгации и неравном кроссинговере, предшествующем редукции плоидности. Реплоиды, отличающиеся измененным соотношением жирных кислот в масле, могут использоваться в качестве исходного материала при селекции растений или в иных целях. На основании вышеизложенного можно сделать вывод о соответствии заявляемого технического решения критерию патентоспособности "ИЗОБРЕТАТЕЛЬСКИЙ УРОВЕНЬ". Заявляемое техническое решение соответствует и критерию патентоспособности "ПРОМЫШЛЕННАЯ ПРИМЕНИМОСТЬ", т.к. оно может быть использовано в сельском хозяйстве, и, кроме того, в описании изобретения будут представлены средства и методы, с помощью которых возможно осуществление технического решения в том виде, как оно охарактеризовано в независимом пункте формулы изобретения. Способ осуществляют следующим образом. Начинают с того, что берут диплоидные сорта сои и переводят их на тетраплоидный уровень колхицинированием или другим известным способом, при этом в первом тетраплоидном поколении (C1) выделяют тетраплоиды по характерным морфологическим признакам и прямым подсчетом хромосом. Выделенные тетраплоиды выращивают (в полевых условиях или в условиях искусственного климата) в течение нескольких поколений. В каждом тетраплоидном поколении отбирают рекомбинированные тетраплоидные формы, явно отличающиеся от базового тетраплоидного морфотипа (по любым количественным и качественным признакам), что свидетельствует о структурных изменениях хромосом одного или обоих составляющих тетраплоид диплоидных геномов. Выделенные рекомбинированные тетраплоидные формы выращивают в течение нескольких поколений отдельными популяциями, выделяя в каждом поколении реплоиды с возвратно кратно-пониженной плоидностью по морфологическим признакам и прямым подсчетом хромосом. Оценивают реплоиды по жирно-кислотному составу масла, выделяя формы, отличающиеся от растений с исходной плоидностью измененным соотношением жирных кислот в масле. Все выделенные реплоиды, отличающиеся измененным жирно-кислотным составом масла, используют в качестве исходного материала в селекционных или иных целях. Пример осуществления способа: В качестве исходных форм были взяты сорта сои Фора и Вилана с 2n=40. Их семена высеяли в полевых условиях и в фазу 1-го тройчатого листа растений перевели на тетраплоидный уровень путем обработки апикальных меристем водным раствором колхицина в концентрации 0,05%. Образующиеся ниже зоны колхицинирования диплоидные побеги удаляли по мере их появления. Колхицинированные побеги доводили до созревания и на них убирали семена C1. Полученные семена C1 маркировали и индивидуально проращивали в чашках Петри до получения корешков длиной 2-5 см, отрезали кончик корешка длиной 0,5 см и цитологически исследовали стандартными методиками. Проростки, у которых была установлена удвоенная плоидность (4n=80), высевали в грунт, формируя по каждому сорту тетраплоидную популяцию C1. Начиная с поколения C1 во внутрисортовых популяциях тетраплоидов сои ежегодно выделяли рекомбинированные тетраплоидные формы, явно отличающиеся от базового тетраплоидного морфотипа по скорости и продолжительности этапов органогенеза, высоте растений, характеру ветвления, форме и числу листочков настоящих листьев, длине черешка, окраске опушения, гипокотиля, венчика цветков, оболочки и рубчика семян. В потомстве рекомбинированных тетраплоидов ежегодно выделяли реплоиды по визуально различимым морфологическим признакам, свойственным сое с исходной базовой плоидностью, и проверяли уровень плоидности реплоидов прямым подсчетом хромосом. Выделенные реплоиды сои сравнивали по комплексу качественных и количественных признаков в сравнении с исходным диплоидным сортом, выделяли формы с измененным соотношением жирных кислот в масле (см. табл.). Анализ жирно-кислотного состава масла у реплоидов сои показал наличие генетических рекомбинаций уровня биохимического синтеза отдельных жирных кислот. У реплоидов сорта Вилана отмечено снижение на 4-5 абс.% содержания олеиновой кислоты с одновременным увеличением на 5 абс.% содержания линолевой и на 2-3 абс.% линоленовой кислот. Жирно-кислотный состав масла у реплоидов низкомасличного сорта Фора отличался более широким полиморфизмом и по содержанию олеиновой кислоты варьировал от 18,02 до 46,50% при исходном уровне 22,75%; по содержанию линолевой кислоты от 38,26 до 63,65% при исходном содержании 55,77% и по содержанию линоленовой кислоты от 3,94 до 8,79% при исходном содержании 8,51%. Жирно-кислотный состав соевого масла реплоидных образцов 3584, 3620/1 и 1338/7 (см табл.) с пониженным до 20% и менее содержанием олеиновой кислоты и повышенным до 60% и выше содержанием линолевой кислоты близок к составу жирных кислот в масле дикорастущих предков сои и отличается повышенной F-витаминной активностью. Жирно-кислотный состав масла реплоидных образцов 1338/6 и 1338/1 (см табл. ) с содержанием олеиновой кислоты 40,37 и 41,44% соответственно, практически удвоенным по сравнению с исходным сортом Фора и с пониженным до 43,07 и 40,75% соответственно содержанием линолевой кислоты, являются жирно-кислотными аналогами кунжутного масла. Образец 1338/2 (см табл.) с содержанием олеиновой и линолевой жирных кислот 46,50 и 38,26% соответственно по жирно-кислотному составу аналогичен арахисовому маслу. Реплоидные образцы 1338/1, 1338/2 и 1338/6 (см табл.) с измененным жирно-кислотным составом масла практически не неизвестны в естественном генофонде сои и, следовательно, являются примерами нового соотношения жирных кислот в масле сои, созданного заявляемым способом. Выращивание тетраплоидов в течение нескольких поколений и одновременный с этим отбор в каждом поколении рекомбинированных тетраплоидных форм, отличающихся от базового тетраплоидного морфотипа, позволяет выявить и сохранить все визуально различимые морфологические рекомбинации на тетраплоидном уровне, т. е. сформировать популяцию тетраплоидных рекомбинантов для последующего выделения реплоидов. Размножение отобранных рекомбинантных полиплоидных форм, каждой в отдельности, в течение ряда поколений и выделение реплоидных форм обеспечивает повышение частоты встречаемости реплоидов с признаками генетически детерминированных рекомбинаций жирно-кислотного состава масла, образовавшихся на полиплоидном уровне или в процессе деполиплоидизации при редукции плоидности гамет, т. е. позволяет расширить полиморфизм по соотношению жирных кислот в масле на реплоидном уровне без использования мутагенов различной природы. Определение уровня плоидности реплоидов прямым подсчетом хромосом обеспечивает достоверную идентификацию истинных реплоидов с признаками генетической рекомбинации, исключая отнесение к этой категории стабилизированных тетраплоидов и анеуплоидов, также морфологически отличающихся от исходных диплоидных сортов сои, т.е. обеспечивает увеличение полиморфизма по соотношению жирных кислот у истинных реплоидов. Оценка реплоидов сои по жирно-кислотному составу масла с выделением форм, отличающихся от растений с исходной плоидностью измененным соотношением жирных кислот, позволяет выявить и использовать в селекционных или иных целях все перспективные, генетически детерминированные рекомбинации по жирно-кислотному составу масла, т.е. расширить, таким образом, существующий в естественном генофонде культурной сои полиморфизм по соотношению жирных кислот. Таким образом, заявляемым изобретением обеспечивается достижение поставленной цели и решение поставленной задачи. -излучением или раствором нитрозоэтилмочевины в дозе, критической для диплоидной формы, выращивание поколений M1 и M2 в виде популяций, выделение в поколении М2 реверсивных диплоидных растений (по следующим признакам: большая высота растений, раннее колошение, высокая озерненность колоса), использование семян с отобранных растений для закладки линий (а.с. СССР 1519596, А 01 Н 1/04, публ. 1989 г.). Хотя известный способ по названию не совпадает с названием заявляемого нами изобретения, он по сущности своей может быть использован для изменения жирно-кислотного состава масла, если всей совокупности приемов известного способа будет подвергнута масличная культура (например, соя), а среди полученных реверсивных диплоидных растений, оцененных по жирно-кислотному составу масла, будут выделены реверсанты с измененным соотношением жирных кислот. Недостатком известного способа, взятого нами в качестве прототипа, является то, что для создания форм растений с измененным соотношением жирных кислот в масле используются мутанты реверсивно диплоидного происхождения. Использование физических или химических мутагенов в критических дозах (LD50) приводит к модификации хозяйственно ценных признаков за счет нарушения функциональности отдельных локусов, контролирующих их реализацию. При этом в организме увеличивается количество негативных мутаций, оказывающих аддитивно-негативное действие на жизнеспособность организма в целом. Разнообразие реверсивных диплоидов при использовании -излучения или раствора нитрозоэтилмочевины определяется только морфологическими признаками (большая высота растений, раннее колошение, высокая озерненность колоса). Использующиеся в прототипе приемы для увеличения частоты появления реверсивных диплоидов существенно ограничивают возможности для отбора хозяйственно ценных диплоидных реверсантов, поскольку их отбор ведется только после обработки мутагенами в критических дозах (т.е. при летальности не менее 50% особей). При этом часть полезных генных рекомбинаций, в т.ч. и по жирно-кислотному составу масла, образовавшихся на полиплоидном уровне или при реверсии на диплоидный уровень, будет утеряна из-за образования несовместимых с жизнью организма мутаций, вызванных применением критических доз мутагенов, а большинство вновь образовавшихся признаков, в т.ч. и изначально доминантных у исходного сорта или тетраплоида, приобретут рецессивность из-за частичного повреждения и нарушения функциональности мутировавших локусов. В целом, известный способ, выбранный нами в качестве прототипа, существенно ограничивает получение, выявление и последующее сохранение максимально возможного полиморфизма по жирно-кислотному составу масла. Заявляемым изобретением решается задача повышения эффективности способа за счет расширения полиморфизма по соотношению компонентов масла при получении реверсантов (реплоидов) (впервые введенный нами термин) сои. Цель изобретения - создание форм сои с измененным жирно-кислотным составом масла без использования мутагенов различной природы, путем полиплоидизации растений, последующего отбора рекомбинантных полиплоидов и выделения среди них за счет спонтанной редукции плоидности гамет форм с возвратной кратно-пониженной плоидностью - реплоидов. Технический результат достигается тем, что в известном способе, включающем перевод исходных диплоидных сортов на тетраплоидный уровень колхицинированием или другим известным способом, выделение реверсивных диплоидных растений по морфологическим и другим хозяйственно ценным признакам и использование их в качестве исходного материала, согласно изобретению полученные тетраплоиды выращивают в течение нескольких поколений и одновременно с этим в каждом поколении отбирают рекомбинированные тетраплоидные формы, отличающиеся от базового тетраплоидного морфотипа, отобранные формы размножают каждую в отдельности в течение ряда поколений, в каждом из которых выделяют формы с реверсивно кратно-пониженной плоидностью (реплоиды), плоидность которых определяют по морфологическим признакам и прямым подсчетом хромосом, оценивают реплоиды по жирно-кислотному составу масла, выделяя формы, отличающиеся от растений с исходной плоидностью измененным или новым жирно-кислотным составом масла. Причем реплоиды сои выделяют по морфологическим, биохимическим или иным признакам, например выделяют реплоиды сои с повышенным или пониженным содержанием олеиновой, линолевой, линоленовой и других жирных кислот. Сопоставительный анализ заявляемого технического решения с прототипом позволяет сделать вывод, что заявляемый способ отличается от известного наличием нового приема: выделением рекомбинированных тетраплоидов из внутрисортовой популяции тетраплоидов, полученных удвоением плоидности исходных диплоидных сортов и выращиваемых в течение нескольких поколений, а также порядком выполнения известного приема "выделение реверсивных форм": формы с реверсивно кратно-пониженной плоидностью (реплоиды) выделяют из рекомбинированных тетраплоидных форм при их размножении в течение ряда поколений. Таким образом, заявляемый способ соответствует критерию патентоспособности "НОВИЗНА". Исследуя уровень техники в процессе проведения патентного поиска по всем видам сведений, общедоступных в печати, мы обнаружили, что способ создания форм сои с измененным жирно-кислотным составом масла, включающий заявленную нами совокупность признаков, для специалиста в области селекции явным образом не следует из известного на сегодня существующего уровня техники. Нашими исследованиями на протяжении ряда лет выявлено, что в природе с определенной частотой постоянно осуществляется спонтанная полиплоидизация высших растений. Полиплоидизация приводит к образованию поливалентов в профазе I мейоза I. С этим связаны различные нарушения конъюгации и кроссинговера гомологичных хромосом, вызывающие их структурные изменения. Нерегулярное распределение дочерних хромосом в мейозе у "ранних" автополиплоидов приводит к редукции плоидности гамет и образованию анеуплоидов и возвратных диплоидов (реплоидов). У "ранних" автополиплоидов при мультивалентной конъюгации и кроссинговере могут происходить асимметричные обмены гомологичными участками хромосом, сопровождающиеся дупликациями и делециями. Структурные изменения хромосом могут приводить к количественным и качественным изменениям существующих или к образованию новых признаков. Реплоидные формы образуются на основе составляющих полиплоидный геном, диплоидных геномов, включая рекомбинированные наборы хромосом. При этом образовавшиеся реплоиды могут быть гомологичными исходной диплоидной форме или отличаться от нее, сохраняя возникшие на полиплоидном уровне новые признаки или приобретая их при мультивалентной конъюгации и неравном кроссинговере, предшествующем редукции плоидности. Реплоиды, отличающиеся измененным соотношением жирных кислот в масле, могут использоваться в качестве исходного материала при селекции растений или в иных целях. На основании вышеизложенного можно сделать вывод о соответствии заявляемого технического решения критерию патентоспособности "ИЗОБРЕТАТЕЛЬСКИЙ УРОВЕНЬ". Заявляемое техническое решение соответствует и критерию патентоспособности "ПРОМЫШЛЕННАЯ ПРИМЕНИМОСТЬ", т.к. оно может быть использовано в сельском хозяйстве, и, кроме того, в описании изобретения будут представлены средства и методы, с помощью которых возможно осуществление технического решения в том виде, как оно охарактеризовано в независимом пункте формулы изобретения. Способ осуществляют следующим образом. Начинают с того, что берут диплоидные сорта сои и переводят их на тетраплоидный уровень колхицинированием или другим известным способом, при этом в первом тетраплоидном поколении (C1) выделяют тетраплоиды по характерным морфологическим признакам и прямым подсчетом хромосом. Выделенные тетраплоиды выращивают (в полевых условиях или в условиях искусственного климата) в течение нескольких поколений. В каждом тетраплоидном поколении отбирают рекомбинированные тетраплоидные формы, явно отличающиеся от базового тетраплоидного морфотипа (по любым количественным и качественным признакам), что свидетельствует о структурных изменениях хромосом одного или обоих составляющих тетраплоид диплоидных геномов. Выделенные рекомбинированные тетраплоидные формы выращивают в течение нескольких поколений отдельными популяциями, выделяя в каждом поколении реплоиды с возвратно кратно-пониженной плоидностью по морфологическим признакам и прямым подсчетом хромосом. Оценивают реплоиды по жирно-кислотному составу масла, выделяя формы, отличающиеся от растений с исходной плоидностью измененным соотношением жирных кислот в масле. Все выделенные реплоиды, отличающиеся измененным жирно-кислотным составом масла, используют в качестве исходного материала в селекционных или иных целях. Пример осуществления способа: В качестве исходных форм были взяты сорта сои Фора и Вилана с 2n=40. Их семена высеяли в полевых условиях и в фазу 1-го тройчатого листа растений перевели на тетраплоидный уровень путем обработки апикальных меристем водным раствором колхицина в концентрации 0,05%. Образующиеся ниже зоны колхицинирования диплоидные побеги удаляли по мере их появления. Колхицинированные побеги доводили до созревания и на них убирали семена C1. Полученные семена C1 маркировали и индивидуально проращивали в чашках Петри до получения корешков длиной 2-5 см, отрезали кончик корешка длиной 0,5 см и цитологически исследовали стандартными методиками. Проростки, у которых была установлена удвоенная плоидность (4n=80), высевали в грунт, формируя по каждому сорту тетраплоидную популяцию C1. Начиная с поколения C1 во внутрисортовых популяциях тетраплоидов сои ежегодно выделяли рекомбинированные тетраплоидные формы, явно отличающиеся от базового тетраплоидного морфотипа по скорости и продолжительности этапов органогенеза, высоте растений, характеру ветвления, форме и числу листочков настоящих листьев, длине черешка, окраске опушения, гипокотиля, венчика цветков, оболочки и рубчика семян. В потомстве рекомбинированных тетраплоидов ежегодно выделяли реплоиды по визуально различимым морфологическим признакам, свойственным сое с исходной базовой плоидностью, и проверяли уровень плоидности реплоидов прямым подсчетом хромосом. Выделенные реплоиды сои сравнивали по комплексу качественных и количественных признаков в сравнении с исходным диплоидным сортом, выделяли формы с измененным соотношением жирных кислот в масле (см. табл.). Анализ жирно-кислотного состава масла у реплоидов сои показал наличие генетических рекомбинаций уровня биохимического синтеза отдельных жирных кислот. У реплоидов сорта Вилана отмечено снижение на 4-5 абс.% содержания олеиновой кислоты с одновременным увеличением на 5 абс.% содержания линолевой и на 2-3 абс.% линоленовой кислот. Жирно-кислотный состав масла у реплоидов низкомасличного сорта Фора отличался более широким полиморфизмом и по содержанию олеиновой кислоты варьировал от 18,02 до 46,50% при исходном уровне 22,75%; по содержанию линолевой кислоты от 38,26 до 63,65% при исходном содержании 55,77% и по содержанию линоленовой кислоты от 3,94 до 8,79% при исходном содержании 8,51%. Жирно-кислотный состав соевого масла реплоидных образцов 3584, 3620/1 и 1338/7 (см табл.) с пониженным до 20% и менее содержанием олеиновой кислоты и повышенным до 60% и выше содержанием линолевой кислоты близок к составу жирных кислот в масле дикорастущих предков сои и отличается повышенной F-витаминной активностью. Жирно-кислотный состав масла реплоидных образцов 1338/6 и 1338/1 (см табл. ) с содержанием олеиновой кислоты 40,37 и 41,44% соответственно, практически удвоенным по сравнению с исходным сортом Фора и с пониженным до 43,07 и 40,75% соответственно содержанием линолевой кислоты, являются жирно-кислотными аналогами кунжутного масла. Образец 1338/2 (см табл.) с содержанием олеиновой и линолевой жирных кислот 46,50 и 38,26% соответственно по жирно-кислотному составу аналогичен арахисовому маслу. Реплоидные образцы 1338/1, 1338/2 и 1338/6 (см табл.) с измененным жирно-кислотным составом масла практически не неизвестны в естественном генофонде сои и, следовательно, являются примерами нового соотношения жирных кислот в масле сои, созданного заявляемым способом. Выращивание тетраплоидов в течение нескольких поколений и одновременный с этим отбор в каждом поколении рекомбинированных тетраплоидных форм, отличающихся от базового тетраплоидного морфотипа, позволяет выявить и сохранить все визуально различимые морфологические рекомбинации на тетраплоидном уровне, т. е. сформировать популяцию тетраплоидных рекомбинантов для последующего выделения реплоидов. Размножение отобранных рекомбинантных полиплоидных форм, каждой в отдельности, в течение ряда поколений и выделение реплоидных форм обеспечивает повышение частоты встречаемости реплоидов с признаками генетически детерминированных рекомбинаций жирно-кислотного состава масла, образовавшихся на полиплоидном уровне или в процессе деполиплоидизации при редукции плоидности гамет, т. е. позволяет расширить полиморфизм по соотношению жирных кислот в масле на реплоидном уровне без использования мутагенов различной природы. Определение уровня плоидности реплоидов прямым подсчетом хромосом обеспечивает достоверную идентификацию истинных реплоидов с признаками генетической рекомбинации, исключая отнесение к этой категории стабилизированных тетраплоидов и анеуплоидов, также морфологически отличающихся от исходных диплоидных сортов сои, т.е. обеспечивает увеличение полиморфизма по соотношению жирных кислот у истинных реплоидов. Оценка реплоидов сои по жирно-кислотному составу масла с выделением форм, отличающихся от растений с исходной плоидностью измененным соотношением жирных кислот, позволяет выявить и использовать в селекционных или иных целях все перспективные, генетически детерминированные рекомбинации по жирно-кислотному составу масла, т.е. расширить, таким образом, существующий в естественном генофонде культурной сои полиморфизм по соотношению жирных кислот. Таким образом, заявляемым изобретением обеспечивается достижение поставленной цели и решение поставленной задачи.
Формула изобретения1. Способ создания форм сои с измененным жирно-кислотным составом масла, включающий перевод исходных диплоидных сортов на тетраплоидный уровень колхицинированием или другим известным способом, выделение реверсивных диплоидных растений по морфологическим и другим хозяйственно ценным признакам и использование их в качестве исходного материала, отличающийся тем, что полученные тетраплоиды выращивают в течение нескольких поколений и одновременно с этим в каждом поколении отбирают рекомбинированные тетраплоидные формы, отличающиеся от базового тетраплоидного морфотипа, отобранные формы размножают, каждую в отдельности, в течение ряда поколений, в каждом из которых выделяют формы с реверсивно кратно-пониженной плоидностью (реплоиды), плоидность которых определяют по морфологическим признакам и прямым подсчетам хромосом, оценивают реплоиды по жирно-кислотному составу масла, выделяя формы, отличающиеся от растений с исходной плоидностью измененным или новым жирно-кислотным составом масла. 2. Способ по п. 1, отличающийся тем, что реплоиды сои выделяют по морфологическим, биохимическим или иным признакам. 3. Способ по п. 2, отличающийся тем, что выделяют реплоиды сои с повышенным или пониженным содержанием олеиновой, линолевой, линоленовой и других жирных кислот.MM4A Досрочное прекращение действия патента из-за неуплаты в установленный срок пошлины за поддержание патента в силе Дата прекращения действия патента: 04.01.2009 Дата публикации: 20.05.2011 Популярные патенты: 2264065 Способ возделывания сельскохозяйственных культур на корм ... (табл.1). Таблица 1Влияние способа посева ярового ячменя с бобовой культурой на динамику урожайности абсолютно сухого вещества, ц/га Способ посева ярового ячменяФаза развития ярового ячменявыход в трубкуколошениемолочное состояниетестообразное состояние восковая спелостьОдновидовой 22,030,0 43,657,749,7 Известный (в смеси с горохом) 23,933,748,2 63,556,3 Предложенный (в смеси с донником) 20,930,546,9 63,259,5 При этом урожайность ячменно-гороховой и ячменно-донниковой смеси практически одинакова. Не отличались они и по качеству получаемого корма. В 1 кг сухого вещества ячменно-гороховой смеси содержалось 9,83 МДж обменной энергии и 10,68% сырого протеина, в ... 2262826 Способ сташевского и.и. переработки навоза личинками синантропных мух и устройство для его осуществления ... ветеринарными свидетельствами, справками. Все поголовье в период профилактического карантина вакцинируют против инфекционных заболеваний в зависимости от эпизоотической обстановки данной местности. Выборочно исследуют свиней и птицу на наличие гельминтов. Проводят профилактические прививки и обработку против гельминтов. Своевременно проводят уборку и утилизацию трупов и борьбу с грызунами и паразитами. Для удаления помета кур замыкают электрическую цепь, питающую электрический двигатель 41. Электрический двигатель 41 приводит в движение цепь 39. Цепь 39 перемещает скребки 40. Скребки 40 захватывают и перемещают куриный помет из под насестов 21 через карантинную зону 18. В ... 2434381 Технологическая линия для приготовления и раздачи влажных кормов ... открытия-закрытия заслонки 28 предусмотрен клапан электромагнитный 30. Заслонка 28 установлена на образующей дозатора 8 ниже электромагнитного клапана 30. Электромагнитный клапан 30 установлен на образующей дозатора 8. На электромагнитном клапане 30 установлен таймер 31. Для установки времени открытия-закрытия заслонки 28 предусмотрен таймер 31. Спиральный трубопровод 29 предназначен для уменьшения скорости истечения сухого комбикорма из дозатора 8. Под кольцевым трубопроводом 17 выполнена кормушка 6.На (фиг.4) представлено дозирующее устройство, состоящее из трубопровода 16, крана 32, телескопического рычага с шаром 33, вилки 34. Электромагнитный клапан 35 (фиг.2, 3) предназначен ... 2141182 Культиватор ... балки 37 и ось соединения поводков 20-24 в кронштейнах 19 рамы 1 обеспечивают копирование рельефа поля и устойчивое выполнение технологического процесса механического уничтожения сорняков. Влажность почвы в верхнем ее слое не оказывает существенного влияния на выполнение технологического процесса - механического подавления сорной растительности. При демонтаже сменных секций 7-11 рабочих органов с катками 25 и барабанами 26 с рамы 1, культиватор можно использовать со стрельчатыми лапами шириной захвата 270 и 330 мм как паровой культиватор для обработки незасоренных полей. Изложенные сведения свидетельствуют о возможности выполнения заявленного изобретения. ФОРМУЛА ... 2496298 Узел крепления пальцев подборщика ... вероятность захватывания стеблей крепежными элементами. Хотя и не исключает ее полностью, поскольку существует возможность захвата стеблей гайкой и выступающей частью болта. Другим недостатком прототипа является трудоемкость сборочно-разборочных операций при обслуживании подборщика, в частности то, что для монтажа и демонтажа пальцев требуется свинчивание и навинчивание гаек на болт и удержание шайб. Кроме того, в процессе эксплуатации подборщика вследствие переменных нагрузок и вибрации возможно ослабление болтового соединения и нарушение прочности крепления пальцев на кронштейнах, что приводит к выпадению пальцев, поломкам подборщика и нарушению технологического процесса ... |
Еще из этого раздела: 2264075 Рулонный пресс-подборщик лубяных культур 2248352 Замещенные бензоилциклогександионы, гербицидное средство на их основе, исходное соединение 2091380 Производные пиколиновой кислоты или их кислотно-аддитивные соли, способ их получения, нербицидная композиция и способ борьбы с сорняками 2263431 Устройство для предпосевной обработки семян 2076603 Способ повышения урожайности сельскохозяйственных культур 2106082 Устройство для укладки подстилочного навоза в бурт 2474105 Плодосъемник шолина 2414114 Зерноуборочный комбайн 2150193 Установка для бесфреонового охлаждения молока 2084104 Ручная сеялка для разбросного посева семян травосмесей |
Изобретения в сельском хозяйстве
Обработка почвы в сельском и лесном хозяйствах
Посадка, посев, удобрение
Уборка урожая, жатва
Обработка и хранение продуктов полеводства и садоводства
Садоводство, разведение овощей, цветов, риса, фруктов, винограда, лесное хозяйство
Новые виды растений или способы их выращивания
Производство молочных продуктов
Животноводство, разведение и содержание птицы, рыбы, насекомых, рыбоводство, рыболовство
Поимка, отлов или отпугивание животных
Консервирование туш животных, или растений или их частей
Биоцидная, репеллентная, аттрактантная или регулирующая рост растений активность химических соединений или препаратов
Хлебопекарные печи, машины и прочее оборудование для хлебопечения
Машины или оборудование для приготовления или обработки теста
Обработка муки или теста для выпечки, способы выпечки, мучные изделия
|
|
||