Высокомасличный устойчивый к имидазолинону подсолнечникПатент на изобретение №: 2420059 Автор: ГЕРДЕС Джеймс Т. (US), БЕНСОН Роберт М. (US), ПОСИ ХАУРЕГИ Гильермо Н. (AR), ЛОПЕС ОЛАСИРЕГИ Мария М. (AR) Патентообладатель: ДАУ АГРОСАЙЕНСИЗ ЛЛС (US) Дата публикации: 10 Ноября, 2009 Начало действия патента: 28 Сентября, 2006 Адрес для переписки: 129090, Москва, ул. Б.Спасская, 25, стр.3, ООО "Юридическая фирма Городисский и Партнеры", пат.пов. Е.Е.Назиной Сорта подсолнечника OI1601А, OI2653R, OI1601В и Е83329, обладающие высоким содержанием олеиновой кислоты и устойчивостью к имидазолинону, продуцирующего семена, депонированные в АТСС под депозитными номерами РТА-9470, РТА-9472, РТА-9471 и РТА-9473, соответственно, скрещивают с иным инбредным растением. Собранные гибридные семена подсолнечника имеют содержание олеиновой кислоты более 80% по массе. 2 з.п. ф-лы, 16 табл. Эта заявка испрашивает приоритет предварительной заявки США No.60/721181, поданной 28 сентября 2005. Это изобретение относится к новому растению подсолнечника (Helianthus sp .), к продуктам, получаемым из нового растения, и способам получения продуктов из подсолнечника. Подсолнечник является одним из немногих сельскохозяйственных видов, происходящих из Северной Америки. Возможно, он был культивирован коренными американскими племенами около 1000 лет до нашей эры. Первые европейцы заметили выращивание подсолнечника во многих местах по всей Северной Америке, от южной Канады до Мексики. Подсолнечник был, возможно, впервые введен в Европу через Испанию, со временем, достигнув России, где он стал широко культивируемым. Селекция для высокой масличности началась в 1860 и привела к увеличению содержания масла с 28 процентов до 50 процентов. Эти высокомасличные линии из России были введены в Соединенные Штаты после II мировой войны. Позднее открытие системы генов, отвечающих за мужскую стерильность и восстановление, сделало возможными гибриды и увеличило коммерческий интерес к урожаю. Впоследствии производство подсолнечников резко возросло в Великих Равнинных штатах, поскольку торговцы находили новые ниши для семян в качестве масличных культур, корма для птиц и в качестве закусочного пищевого продукта для человека. Культивируемый подсолнечник (Helianthus annuus L.) является главным мировым источником растительного масла. В Соединенных Штатах главными производящими подсолнечник штатами являются Дакота, Миннесота, Канзас, Колорадо, Небраска, Техас и Калифорния, несмотря на то, что большинство штатов имеют некоторые коммерческие посевные площади. Производство подсолнечного масла в Соединенных Штатах составило 2,26 миллиона фунтов в 2003. Немасличное производство составило 406000 фунтов. Немасличные подсолнечники составили в среднем 1256 фунтов на акр в 2003, в то время как масличные подсолнечники давали средний урожай 1206 фунтов на акр в 2003. Масличные семена подсолнечников рассматриваются наряду с семенами хлопчатника, соевыми бобами и канолой, а выращивание подсолнечника, как урожай масличной культуры, составило конкуренцию для соевых бобов. Масло составляет 80% от стоимости урожая подсолнечника, по сравнению с соевыми бобами, которые получают большую часть стоимости от пищевых продуктов. Подсолнечное масло повсеместно рассматривается как высококачественное масло из-за своего светлого цвета, высокого уровня ненасыщенных жирных кислот, малого содержания линоленовой кислоты, слабовыраженного вкуса и высоких температур образования копоти. Первичными жирными кислотами в масле являются олеиновая и линолевая, остальные состоят из пальмитиновой и стеариновой насыщенных жирных кислот. Нелущеный или частично лущеный корм успешно заменил бобовый корм в изоазотистых (с одинаковым содержанием белка) диетах для жвачных животных, так же как и для кормления свиней и домашних птиц. В подсолнечном корме выше содержание волокон, ниже энергетическая ценность и ниже содержание лизина, но выше метионина, чем в кормах из соевых бобов. Процент белка в подсолнечном корме изменяется в пределах от 28 процентов для нелущеных семян до 42 процентов для полностью лущеных семян. В дополнение к его применению в пищу и пищевых продуктах для людей и животных подсолнечное масло также имеет промышленное применение. Оно было применено в красках, лаках и пластмассах из-за хороших полувысыхающих свойств, без изменений цвета, связанного с маслами с высоким содержанием линоленовой кислоты. Оно также было применено в производстве мыл, детергентов и косметики. Изучали применение подсолнечного масла (и других растительных масел) в качестве носителей для пестицидов и в производстве химических препаратов, поверхностно-активных веществ, адгезивов, смягчителей тканей, лубрикантов и покрытий. Значительная работа также была сделана для изучения возможности использования подсолнечника в качестве альтернативного источника топлива в дизельных двигателях, поскольку подсолнечное масло обладает 93 процентами энергии дизельного топлива США Номер 2 (октановое число 37). Позднее подсолнечное масло было предложено в качестве источника водорода для водородных топливных элементов (BBC News, август 26, 2004). Подсолнечник является однолетним, прямостоячим, широколистным растением с мощным стержневым корнем и обильным боковым простиранием поверхностных корней. Стебли обычно в начале сезона округлые, угловатые и с поздним одревеснением в сезоне и, как правило, без ветвей. Корзинка подсолнечника представляет собой не один цветок (как подразумевает название), но составлена из 1000 до 2000 индивидуальных цветков, соединенных общим ложем. Цветки на окружности являются язычковыми цветками без тычинок и пестиков; остальные цветки являются совершенными цветками с тычинками и пестиками. Цветение (сбрасывание пыльцы) начинается с периферии и продолжается к центру корзинки. Поскольку многие сорта подсолнечника имеют степень самонесовместимости, перенос пыльцы насекомыми между растениями важен, а семьи пчел, как правило, увеличивают урожай. Развитие цитоплазматической системы, отвечающей за мужскую стерильность и восстановление для подсолнечника, позволило семенным компаниям производить высококачественные гибридные семена. Большинство из них дают большие урожаи, чем перекрестноопыляющиеся сорта, и содержат более высокий процент масла. Продуктивность сортов, тестированных в нескольких условиях окружающей среды, является лучшей основой для выбора гибридов подсолнечника. Выбор должен учитывать урожай, процент масла, зрелость, размер семени (для немасличных рынков) и устойчивость к полеганию и заболеваниям. Урожаи подсолнечника как сельскохозяйственной культуры уменьшаются, а изредка уничтожаются сорняками, которые конкурируют с подсолнечником за влагу и питательные вещества и иногда за свет. Подсолнечник является сильным конкурентом для сорняков, особенно за свет, но не покрывает землю достаточно рано, чтобы предотвратить образование сорняков. По этой причине борьба с сорняками в начале сезона необходима для хороших урожаев; успешная борьба с сорняками должна включать комбинацию агротехнических и химических приемов. Почти все посевы подсолнечника в Северной Америке обрабатываются и/или боронуются для борьбы с сорняками и около 2/3 из них обрабатывается гербицидами. Имидазолиноны представляют собой класс гербицидов, которые действуют против широкого спектра сорняков при низких концентрациях и применяются по всему миру в бобовых, зерновых насаждениях и плантационных культурах. Эти гербициды широко применяются не только из-за их эффективности, но также из-за их низкой токсичности для млекопитающих и слабого воздействия на окружающую среду. Наличие устойчивых к имидазолинону сельскохозяйственных культур дает много пользы и преимуществ плантаторам, обеспечивая разработку очень гибкой программы по борьбе с сорняками. По причине широкого спектра активности и гибких методов внесения имидазолинонов программа по борьбе с сорняками, которая использует устойчивые сельскохозяйственные культуры, может базироваться на сорняках, бороться с которыми можно с меньшей заботой об относительной селективности гербицида. Культуры, устойчивые к имидазолинону, таким образом, могут служить эффективным средством борьбы с сорняками. Качества коммерчески конкурентоспособных сортов растения, в основном, включают более чем высокую урожайность с превосходной устойчивостью к полеганию. Поскольку урожайность является единственным, наиболее решающим фактором производства, который влияет на прибыль производителя культур, производители ожидают рост урожая от года к году, устойчивости к заболеваниям, других значимых признаков, и более часто - устойчивости к гербицидам. Добавление устойчивости к гербицидам создало как возможности, так и огромные задачи в продуктивном сельском хозяйстве. Ссылки: Putnam, et al. 1990. Sunflower in Alternative Field Crops Manual. University of Minnesota: Center for Alternative Plant & Animal Products; Minnesota Extension Service; Boland, M. and Stroade, J. 2004. Sunflower Industry Profile. Department of Agricultural Economics, Kansas State University; Agricultural Marketing Resource Center; Duke, Stephen, Ed. 1996. Herbicide-Resistant Crops. Agricultural, Environmental, Economic, Regulatory, and Technical Aspects. CRC Press; Патент Соединенных Штатов No. 4627192; Патент Соединенных Штатов No. 5276264; Патент Соединенных Штатов No. 6388113. Вышеуказанные примеры в связанной области техники и ограничения, связанные с ними, подразумеваются иллюстративными, а не исключительными. Другие ограничения в связанной области техники будут понятны специалистам в данной области техники при прочтении описания. Следующие способы осуществления и их аспекты описаны и проиллюстрированы вместе с системами, средствами и способами, которые подразумеваются типичными и иллюстративными, но не ограничивающими объем изобретения. В различных способах осуществления одна или более вышеописанных проблем были уменьшены или исключены, в то время как другие способы осуществления направлены на другие усовершенствования. Аспектом настоящего изобретения является предоставление гибридного семени подсолнечника, которое имеет общее содержание олеиновой кислоты, по меньшей мере, 85,2 процента и обладает устойчивостью к имидазолинонам. Другим аспектом настоящего изобретения является предоставление новых растений подсолнечника, которые могут быть эффективно применены для производства родительских линий и гибридов, обладающих желаемыми агрономическими признаками в комбинации с высоким содержанием олеиновой кислоты. Еще одним аспектом настоящего изобретения является предоставление способа получения гибридного подсолнечника, устойчивого к имидазолинонам. В завершение вышеуказанных аспектов было предоставлено, в соответствии с настоящим изобретением, семя подсолнечника с содержанием олеиновой кислоты более 85,2 процентов и толерантностью к имидазолинону. В соответствии с еще одним аспектом настоящего изобретения был предоставлен сорт подсолнечника с содержанием олеиновой кислоты, по меньшей мере, 85,2 процентов и устойчивостью к имидазолинонам. Другие аспекты, признаки и преимущества настоящего изобретения будут понятны из следующего детального описания. Однако следует понимать, что детальное описание и специфические примеры, при указании предпочтительных способов осуществления изобретения, даются только для иллюстрации, в связи с этим различные изменения и модификации в пределах сущности и объема изобретения будут понятны специалистам в данной области техники из этого детального описания. В следующем описании и примерах в этом документе используется ряд терминов. Чтобы обеспечить ясное и согласующееся понимание спецификации и формулы изобретения, включая объем даваемый в этих терминах, представлены следующие определения: ALS ингибитор. Как применено здесь, ALS ингибитор означает любую гербицидно-эффективную форму сульфонилмочевины, триазолопиримидинсульфонамидов, имидазолинонов или гетероарильных эфиров, включая любую их соль. Аллель. Аллель является любой из одной или более альтернативных форм гена, все аллели которого относятся к одному признаку или характеристике. В диплоидной клетке или организме две аллели данного гена занимают соответствующие локусы на паре гомологичных хромосом. Обратное скрещивание. Обратное скрещивание является процессом, при котором селекционер многократно скрещивает гибридное потомство с одним из родителей, например, первое поколение гибридов F1 с одним из родительских генотипов F1 гибрида. Коммерчески приемлемый. Термин коммерчески приемлемый означает сорт подсолнечника или гибрид, дающий урожай зерна более 2000 фунтов на акр, по меньшей мере, в течение 2 лет и в 10 средах обитания. FAME анализ. Анализ Жирнокислотных Метиловых Эфиров (FAME) является способом, который обеспечивает точное количественное определение жирных кислот, которые составляют классы сложных липидов. Устойчивость к имидазолинону (Imi). Устойчивость и/или толерантность придается одним или более генами, которые изменяют ацетолактатсинтазу (ALS), также известную, как синтаза ацетогидроксикислоты (AHAS), позволяя ферменту противостоять действию имидазолинона. Содержание масла. Содержание масла измеряется как процент от массы всего сухого семени и является характеристичным для разных сортов. Оно может быть определено, используя разные аналитические методы, такие как ЯМР, спектрометрия в ближней ИК-области и экстракция в аппарате Сокслета. Содержание олеиновой кислоты. Олеиновая кислота является мононасыщенной жирной кислотой с химической формулой С18 Н34О2. Ее название по ИЮПАК - цис-9-октадециленовая кислота, но обычно она именуется как С18:1. Содержанием олеиновой кислоты называется процент всей фракции жирных кислот подсолнечного масла, которая состоит из С18:1. Процент олеиновой кислоты (OLE). Процент масла семени, которое является олеиновой кислотой. Процент всех жирных кислот. Процент всех жирных кислот определяется путем экстрагирования образца масла из семени, получения сложных метиловых эфиров жирных кислот, присутствующих в этом образце масла, и анализа пропорций различных жирных кислот в образце, используя газовую хроматографию. Состав жирных кислот также может быть отличительной характеристикой сорта. Содержание белка. Содержание белка измеряется как процент всего сухого семени и является характеристичным для разных сортов. Оно может быть определено при помощи разных аналитических методов, таких как ближняя ИК-область и Кьельдаль. Устойчивость к полеганию. Устойчивость к полеганию измеряет способность сорта стоять в поле в условиях высокого урожая и суровых факторов окружающей среды. Сорт может иметь хорошую (остается вертикальным), среднюю или плохую (полегает) устойчивость к полеганию. Степень устойчивости к полеганию не выражается для всех условий, но является наиболее значимой, когда присутствует некоторая степень полегания в полевом испытании. Конвертированный Одиночный Ген (Конверсия). Растение с одиночным конвертированным геном (конверсия) относится к растениям, которые получают методом селекции растений, называемым обратным скрещиванием, при котором существенно все желаемые морфологические и физиологические характеристики сорта получены в дополнение к одиночному гену, перенесенному в сорт посредством способа обратного скрещивания или посредством генной инженерии. Всего Насыщенных (TOTSAT). Общий процент масла насыщенных жиров в масле семени, включая С12:0, С14:0, С16:0, С18:0, С20:0, С22:0 и С24:0. Средний Урожай (урожайность). Средний урожай всех посевов подсолнечника, выращиваемых на данной местности. Урожай. На 10 процентов больше среднего урожая в 10 или более местностях. Контрольное Среднее. Среднее для одного или более контрольных сортов или гибридов на данной местности. До настоящего изобретения никогда не разрабатывался сорт подсолнечника, обладающего вместе как маслом с высоким содержанием олеиновой кислоты, так и устойчивостью к имидазолинону, совмещенными в одном генотипе подсолнечника. Эти признаки не были ранее совмещены ни в одном коммерческом или диком типе подсолнечника. Присутствие обоих признаков в одном сорте подсолнечника существенно расширяет полезность сельскохозяйственной культуры, обеспечивая высокожелаемым маслом с высоким содержанием олеиновой кислоты и большую гибкость в борьбе с сорняками. Все виды сельскохозяйственных культур выращиваются с целью сбора урожая некоторого продукта, имеющего коммерческую значимость. Увеличение продуктивности или урожая такого продукта является главной целью большинства программ по селекции растений. Высшим приоритетом в большинстве программ по разработке сортов подсолнечника является увеличение урожая семян. Урожайность семян является количественной характеристикой, контролируемой многими генами, и сильно зависимым от окружающей среды. Наследуемость урожая является самым низким и самым изменчивым из главных агрономических признаков, рассматриваемых в разработке сортов, с оценочными показателями наследуемости, варьирующими от 3 до 58 процентов. Урожайность является примером количественной характеристики, которую селекционеры пытаются улучшить сверх его настоящего уровня в современных сортах. Устойчивость к заболеваниям требуется в большинстве случаев для защиты урожайного потенциала сорта. Введение устойчивого или толерантного к гербицидам признака в высокоурожайные сорта является трудной задачей. Затруднение увеличивается на несколько порядков, если селекционер пытается совместить устойчивость к гербицидам с высоким содержанием олеиновой кислоты в масле в одном сорте. Растениеводу-селекционеру, чтобы найти сорт с достаточной ценностью (например, высокая урожайность), для увеличения коммерческого сбыта необходимо сделать много скрещиваний и вырастить тысячи экспериментальных генотипов. Оценка такого количества генотипов является большой проблемой и потребляет неимоверное количество времени и средств селекционера. В некоторых случаях с момента первого скрещивания до момента, когда получен коммерчески приемлемый генотип, может пройти десяток лет. Эффективность селекции для генотипов с представляющими интерес признаками (например, высота, урожай, устойчивость к гербицидам, масло с высоким содержанием олеиновой кислоты) в программе селекции будет зависеть от следующего: 1) степень, до которой вариабельность представляющих интерес признаков индивидуальных растений в популяции является результатом генетических факторов и, таким образом, передается потомству выбранных генотипов; и 2) насколько вариабельность представляющих интерес признаков (например, высота, урожай, устойчивость к гербицидам, масло с высоким содержанием олеиновой кислоты) среди растений вызвана окружающей средой, в которой выращиваются разные генотипы. Наследственности признаков изменяются от контролирования одного главного гена, экспрессия которого не зависит от окружающей среды (например, качественные характеристики), до контроля многими генами, эффект от которых зависит от окружающей среды (например, количественные характеристики). Селекция для количественных признаков дополнительно характеризуется тем фактом, что: 1) разницы, получаемые от эффекта каждого гена, являются малыми, делая трудным или невозможным их индивидуальную идентификацию; 2) количество генов, вносящих вклад в характеристику, настолько велико, что отчетливые сегрегационные отношения редко, если вообще когда-либо, получают; и 3) эффекты генов могут быть выражены разными способами, основываясь на изменениях под воздействием внешней среды. Таким образом, точная идентификация трансгрессивных сегрегантов и первосортных генотипов с интересующими признаками является крайне трудной, и ее успешность зависит от возможности растениевода-селекционера минимизировать влияние изменений окружающей среды на выражение количественной характеристики в популяции. Вероятность идентификации трансгрессивного сегреганта сильно сокращается с увеличением количества признаков, совмещаемых в одном генотипе. Например, если скрещивание производится между сортами, различающимися в трех сложных характеристиках, таких как урожайность, устойчивость к гербицидам и высокое содержание олеиновой кислоты в масле, крайне сложно получить одновременно путем рекомбинации максимальное количество благоприятных генов для каждой из трех характеристик в одном генотипе. Следовательно, все, на что может надеяться селекционер - это получение благоприятного ассортимента генов для первой сложной характеристики, совмещенного с благоприятным ассортиментом генов для второй характеристики в одном генотипе в дополнение к гену устойчивости к гербицидам. Способы, применяемые в программах разработки сортов, и вероятность их успеха зависят от количества улучшаемых одновременно характеристик, таких как признаки урожайности семян, устойчивости к заболеваниям и устойчивости/толерантности к гербицидам. Пропорцию желаемых индивидуумов с множеством характеристик в популяции получают путем перемножения пропорции желаемых индивидуумов, ожидаемых в популяции, для каждой улучшаемой характеристики. Это предполагает, что характеристики наследуются независимо, т.е. не связаны генетически. Эти принципы могут быть применены не только к традиционно скрещиваемым линиям, но также к линиям, содержащим один или более трансгенов. Как при комбинировании желаемых традиционных или трансгенных признаков посредством скрещивания трансгенных линий, так и при котрансформации множества генов в одну линию, совместный эффект на урожай может быть мультипликативным. Вероятность идентификации линии с подходящей комбинацией признаков дополнительно уменьшается, когда рассматриваются потенциальные эффекты трансгена на регуляцию метаболизма в растении. Например, можно рассматривать потенциальный эффект генов, отвечающих за устойчивость к имидазолинонам. Ген, отвечающий за этот признак, является геном, кодирующим мутированный фермент ацетолактатсинтазу (ALS). ALS ген влияет на тесно связанные биохимические реакции при синтезе аминокислот. Приемлемые линии имеют фоновые генотипы, которые компенсируют или, главным образом, не изменяются при пертурбациях, вызываемых введенным геном. Когда линии с приемлемой устойчивостью к гербицидам комбинируют селекцией с линиями с высоким содержанием олеиновой кислоты в масле, фоновые генотипы, которые приспособились к введенным или мутированным генам, комбинируются и становится необходимым выбирать новые генотипы. Частота генотипов с подходящим урожаем будет соответственно сокращена. В силу чего совмещение устойчивости к гербицидам с высоким урожаем и высоким содержанием олеиновой кислоты в данном сорте подсолнечника или гибрида является крайне трудным препятствием. Неожиданно, признаки устойчивости к имидазолинону с высоким содержанием олеиновой кислоты были совмещены в коммерчески приемлемом сорте настоящего изобретения. Будучи однажды совмещенными в сорте, эти признаки далее могут быть перенесены на другие генетические фоны. ПРИМЕРЫ Следующие примеры представлены для дополнительной иллюстрации настоящего изобретения и не предполагают ограничение изобретения за пределами, предложенными в прилагающихся пунктах формулы изобретения. Пример 1 Гибрид подсолнечника Е83329, имеющий высокое содержание олеиновой кислоты и устойчивость к имидазолинону. Одним примером устойчивости к имидазолинону и высокого содержания олеиновой кислоты является сорт подсолнечника Е83329. Е83329 был создан посредством селекции растений и является устойчивым и однообразным. Этот подсолнечник с высоким содержанием олеиновой кислоты, который является устойчивым к имидазолинонам. Некоторые из критериев, использованных при выборе в разных поколениях, включают: урожай семян, устойчивость к полеганию, прорастание, толерантность к заболеваниям и зрелость. Гибрид показал однообразие и устойчивость, как описано в информации для следующего описания сорта. Родительские линии самоопылялись достаточное количество поколений с тщательным вниманием к однообразию типа растения. Гибрид размножали с непрерывным наблюдением за однообразием. Е83329 обладает следующими морфологическими и другими характеристиками (см. таблицу 1): Таблица 1 Растение: Высота: 80 дюймов Количество листьев: 28 Форма листа: Сердцевидная Длина листа: 11 дюймов Ширина листа: 10,7 дюймов Изрезанность края листа: Средняя Положение листа: Поникающее Дней до цветения: 68 Дней до созревания: 98 Цвет язычкового цветка: Желтый Летучки: Зеленые Диаметр корзинки: 7 дюймов Форма корзинки: выпуклая Положение корзинки: поникающее Количество семян на корзинку: 1675 Масса семени (г/200): 10 Влажность урожая (процент): 10,6 Урожай (фунт/акр): 2910 Процент масла: 41 Состав масла: Олеиновая: 86,9 процента С16:0: 4,05 процента С16:1: 0,18 процента С18:0: 2,93 процента С18:2: 3,01 процента Насыщенные: 8,89 процента Устойчивость к имидазолинону: Отличная Растение: Высота: 80 дюймов Количество листьев: 28 Форма листа: Сердцевидная Длина листа: 11 дюймов Ширина листа: 10,7 дюймов Изрезанность края листа: Средняя Положение листа: Поникающее Дней до цветения: 68 Дней до созревания: 98 Цвет язычкового цветка: Желтый Летучки: Зеленые Диаметр корзинки: 7 дюймов Форма корзинки: выпуклая Положение корзинки: поникающее Количество семян на корзинку: 1675 Масса семени (г/200): 10 Влажность урожая (процент): 10,6 Урожай (фунт/акр): 2910 Процент масла: 41 Состав масла: Олеиновая: 86,9 процента С16:0: 4,05 процента С16:1: 0,18 процента С18:0: 2,93 процента С18:2: 3,01 процента Насыщенные: 8,89 процента Устойчивость к имидазолинону: Отличная В Таблице 2, которая следует ниже, выбранные характеристики Е83329 сравниваются с коммерческим сортом 7350. Таблица 2 Признак Е833297350 Дней до цветения68 64 Дней до созревания 9896 Высота 80 дюймов73 дюйма Количество листьев28 24Диаметр корзинки7 дюймов 8 дюймов Кол-во семян на корзинку 1675 1822Масса семени (г/200)10 10 Урожай (фунтов/акр) 29103034В Таблице 3, которая следует ниже, сравниваются составы масел из Е83329 и коммерческого сорта 7350. Таблица 3 Признак масла Е83329 7350 Общий процент масла 41 процент45,2 процента Процент олеиновой 86,987,4 Процент С16:0 4,05 3,77Процент С16:10,18 0,15Процент С18:02,93 3,11Процент С18:23,01 2,93Процент насыщенных8,89 8,62В Таблице 4, которая следует ниже, сравниваются устойчивости к имидазолинону Е83329 и коммерческого сорта 7350 по шкале от 1 до 9, где 1 - отличная устойчивость, а 9 - плохая устойчивость. Столбец 1 показывает дозу применяемого гербицида и время после применения, когда была измерена устойчивость. IMI - имидазолиноновый гербицид, 1X IMI - однократная стандартная доза имидазолинона и так далее. Таблица 4 Устойчивость к имидазолинону Е83329 7350 1 неделя после опрыскивания 1X IMI2,0 9,02X IMI3,0 9,03X IMI3,5 9,03 недели после опрыскивания 1X IMI1,0 9,02X IMI1,0 9,03X IMI1,0 9,0В Таблице 5, которая следует ниже, сравниваются данные FAME анализов для Е83329 и коммерческого сорта 7350. Каждое значение представляет процент от общего масла жирных кислот. Таблица 5 Состав Масла Е83329 7350 С14:00,06 0,05С16:0 4,05 3,77С16:1 0,18 0,15С18:0 2,93 3,11С18:1 86,90 87,42С18:2 3,01 2,93С18:3 0,09 0,08С20:0 0,35 0,34С20:1 0,32 0,33С20:2 0,01 0,01С22:0 1,10 0,98С24:0 0,40 0,36С24:1 0,01 0,01ИТОГО 8,89 8,62Пример 2 Сорт подсолнечника OI1601A, обладающий устойчивостью к имидазолинону и высоким содержанием олеиновой кислоты. Вторым примером устойчивости к имидазолинону и высокого содержания олеиновой кислоты является сорт подсолнечника OI1601A. OI1601A был разработан посредством селекции растений и является устойчивым и однообразным. Этот подсолнечник с высоким содержанием олеиновой кислоты, который является устойчивым к имидазолинонам. Некоторые из критериев, использованных при выборе в разных поколениях, включают: урожай семян, устойчивость к полеганию, прорастание, толерантность к заболеваниям и зрелость. Сорт показал однообразие и устойчивость, как описано в информации для следующего описания сорта. Он самоопылялся достаточное количество поколений с тщательным вниманием к однообразию типа растения. Сорт увеличивали с непрерывным наблюдением за однообразием. OI1601A обладает следующими морфологическими и другими характеристиками (см. таблицу 6): Таблица 6 Растение: Высота: 57 дюймов Количество листьев: 29 Форма листа: Сердцевидная Длина листа: 10,6 дюймов Ширина листа: 10,2 дюймов Изрезанность края листа: Средняя Положение листа: Поникающее Дней до цветения: 68 Дней до созревания: 95 Цвет язычкового цветка: Желтый Летучки: Зеленые Диаметр корзинки: 7 дюймов Форма корзинки: выпуклая Положение корзинки: поникающее Процент масла: 40,9 Состав масла: Олеиновая: 88,95 процента С16:0: 3,53 процента С16:1: 0,13 процента С18:0: 2,49 процента С18:2: 2,98 процента Насыщенные: 7,55 процента Устойчивость к имидазолинону: Отличная В Таблице 7, которая следует ниже, выбранные характеристики OI1601A сравниваются с коммерческим сортом 7350. Таблица 7 Признак OI1601A7350 Дней до цветения68 64 Дней до созревания 9596 Высота 57 дюймов73 дюйма Количество листьев29 24Диаметр корзинки7 дюймов 8 дюймов Кол-во семян на корзинку 463 1822Масса семени (г/200)12 10 Урожай (фунтов/акр) 11253034В Таблице 8, которая следует ниже, сравниваются составы масел из OI1601A и коммерческого сорта 7350. Таблица 8 Признак масла OI1601A 7350 Общий процент масла 40,945,2 процента Процент Олеиновой 88,95 87,4Процент С16:03,53 3,77Процент С16:10,13 0,15Процент С18:02,49 3,11Процент С18:22,98 2,93Процент насыщенных7,55 8,62В Таблице 9, которая следует ниже, сравниваются устойчивости к имидазолинону OI1601A и коммерческого сорта 7350 по шкале от 1 до 9, где 1 - отличная устойчивость, а 9 - плохая устойчивость. Столбец 1 показывает дозу применяемого гербицида и время после применения, когда была измерена устойчивость. IMI - имидазолиноновый гербицид, 1X IMI - однократная стандартная доза имидазолинона, и так далее. Таблица 9 Устойчивость к имидазолинону OI1601A 7350 1 неделя после опрыскивания 1X IMI2,0 9,02X IMI3,0 9,03X IMI3,5 9,03 недели после опрыскивания 1X IMI1,0 9,02X IMI1,0 9,03X IMI1,0 9,0Пример 3 Сорт подсолнечника OI2653R, имеющий высокое содержание олеиновой кислоты и устойчивость к имидазолинону. Третьим примером устойчивости к имидазолинону и высокого содержания олеиновой кислоты является сорт подсолнечника OI2653R. OI2653R был создан посредством селекции растений и является устойчивым и однообразным. Этот подсолнечник с высоким содержанием олеиновой кислоты, который является устойчивым к имидазолинонам. Некоторые из критериев, использованных при выборе в разных поколениях, включают: урожай семян, устойчивость к полеганию, прорастание, толерантность к заболеваниям и зрелость. Сорт показал однообразие и устойчивость, как описано в информации для следующего описания сорта. Он самоопылялся достаточное количество поколений с тщательным вниманием к однообразию типа растения. Сорт размножали с непрерывным наблюдением за однообразием. OI2653R обладает следующими морфологическими и другими характеристиками (см. таблицу 10): Таблица 10 Растение: Высота: 62 дюймов Количество листьев: 24 Форма листа: Сердцевидная Длина листа: 11,7 дюймов Ширина листа: 9,8 дюймов Изрезанность края листа: Средняя Положение листа: Поникающее Дней до Цветения: 74 Дней до Созревания: 102 Цвет язычкового цветка: Желтый Летучки: Зеленые Диаметр корзинки: 4,5 дюймов Форма корзинки: плоская Положение корзинки: поникающее Процент масла: 42,8 Состав масла: Олеиновая: 89 процента С16:0: 3,4 процента С16:1: 0,11 процента С18:0: 2,26 процента С18:2: 3,26 процента Насыщенные: 7,15 процента Устойчивость к имидазолинону: Отличная В Таблице 11, которая следует ниже, выбранные характеристики OI2653R сравниваются с коммерческим сортом 7350. Таблица 11 Признак OI2653R7350 Дней до цветения74 64 Дней до созревания 10296 Высота 62 дюймов73 дюйма Количество листьев24 24Диаметр корзинки4,5 дюймов 8 дюймов Кол-во семян на корзинку 510 1822Масса семени (г/200)7 10 Урожай (фунтов/акр) 4003034В Таблице 12, которая следует ниже, сравниваются составы масел OI2653R и коммерческого сорта 7350. Таблица 12 Признак масла OI2653R 7350 Общий процент масла 42,8 процента45,2 процента Процент олеиновой 8987,4 Процент С16:0 3,4 3,77Процент С16:10,11 0,15Процент С18:02,26 3,11Процент С18:23,26 2,93Процент насыщенных7,15 8,62В Таблице 13, которая следует ниже, сравниваются устойчивости к имидазолинону OI2653R и коммерческого сорта 7350 по шкале от 1 до 9, где 1 - отличная устойчивость, а 9 - плохая устойчивость. Столбец 1 показывает дозу применяемого гербицида и время после применения, когда была измерена устойчивость. IMI - имидазолиноновый гербицид, 1X IMI - однократная стандартная доза имидазолинона, и так далее. Таблица 13 Устойчивость к имидазолинону OI2653R 7350 1 неделя после опрыскивания 1X IMI2,0 9,02X IMI3,5 9,03X IMI3,5 9,03 недели после опрыскивания 1X IMI1,0 9,02X IMI1,0 9,03X IMI1,0 9,0В Таблице 14, которая следует ниже, сравниваются данные FAME анализов для OI2653R и коммерческого сорта 7350. Каждое значение представляет процент от общего масла жирных кислот. Таблица 14 Состав Масла OI2653R 7350 С14:00,04 0,05С16:0 3,40 3,77С16:1 0,11 0,15С18:0 2,26 3,11С18:1 89 87,42 С18:23,26 2,93С18:3 0,11 0,08С20:0 0,26 0,34Итого 7,15 8,62Пример 4 Сорт подсолнечника OI1601B обладает высоким содержанием олеиновой кислоты и устойчивостью к имидазолинону. Четвертым примером устойчивости к имидазолинону и высокого содержания олеиновой кислоты является сорт подсолнечника OI1601B. OI1601B был создан посредством селекции растений и является устойчивым и однообразным. Этот подсолнечник с высоким содержанием олеиновой кислоты, который является устойчивым к имидазолинонам. Некоторые из критериев, использованных при выборе, в разных поколениях включают: урожай семян, устойчивость к полеганию, прорастание, толерантность к заболеваниям и зрелость. Сорт показал однообразие и устойчивость, как описано в информации следующего описания сорта. Он самоопылялся достаточное количество поколений с тщательным вниманием к однообразию типа растения. Сорт размножали с непрерывным наблюдением за однообразием. OI1601B обладает следующими морфологическими и другими характеристиками (см. таблицу 15): Таблица 15 Растение: Высота: 57 дюймов Количество листьев: 29 Форма листа: Сердцевидная Длина листа: 10,6 дюймов Ширина листа: 10,2 дюймов Изрезанность края листа: Средняя Положение листа: Поникающее Дней до Цветения: 68 Дней до Созревания: 95 Цвет язычкового цветка: Желтый Летучки: Зеленые Диаметр корзинки: 7 дюймов Форма корзинки: выпуклая Положение корзинки: поникающее Количество семян на корзинку: 463 Масса семени (г/200): 12 Влажность урожая (процент): 10 Урожай (фунт/акр): 1125 Процент масла: 40,9 Состав масла: Олеиновая: 88,95 процента С16:0: 3,53 процента С16:1: 0,13 процента С18:0: 2,49 процента С18:2: 2,98 процента Насыщенные: 7,55 процента Устойчивость к имидазолинону: Отличная В Таблице 16, которая следует ниже, сравниваются данные FAME анализов для OI1601B и коммерческого сорта 7350. Каждое значение представляет процент от общего масла жирных кислот. Таблица 16 Состав Масла OI1601B 7350 С14:00,04 0,05С16:0 3,40 3,77С16:1 0,11 0,15С18:0 2,26 3,11С18:1 89 87,42 С18:23,26 2,93С18:3 0,11 0,08С20:0 0,26 0,34ИТОГО 7,15 8,62Это изобретение также направлено на способы получения растения подсолнечника путем скрещивания первого родительского растения подсолнечника со вторым родительским растением подсолнечника, где первое или второе растение подсолнечника является растением подсолнечника из сортов Е83329, OI1601A, OI2653R или OI1601B. Кроме того, оба - первое и второе, родительские растения подсолнечника могут быть из сортов Е83329, OI1601A, OI2653R или OI1601B. Таким образом, любые способы с применением сортов Е83329, OI1601A, OI2653R или OI1601B являются частью этого изобретения: самоопыление, обратное скрещивание, селекция гибридов, скрещивания с популяциями. Любые растения, полученные с применением Е83329, OI1601A, OI2653R или OI1601B в качестве родителя, находятся в объеме этого изобретения. Полезные методы включают, но не ограничиваются указанными, векторы для экспрессии, введенные в растительную ткань посредством использования прямого способа переноса гена, такого как микроинжекторная опосредованная доставка, инъекция ДНК, электропорация и подобные. Более предпочтительно, векторы для экспрессии вводятся в растительные ткани при использовании микроинжекторной доставки биолистическим аппаратом или Agrobacterium-опосредованной трансформации. Трансформированные растения, полученные с протоплазмой по изобретению, подразумеваются входящими в объем этого изобретения. С появлением методов молекулярной биологии, которая сделала возможным выделение и характеризацию генов, которые кодируют специфические белковые продукты, ученые в области биологии растений развивают постоянный интерес к инженерии генома растений для включения и экспрессии чужеродных генов, или дополнительных или модифицированных версий собственных или эндогенных генов (вероятно, управляемых разными промоторами), чтобы изменить признаки растения специфическим образом. Такие чужеродные дополнительные и/или модифицированные гены относятся к собирательному термину «трансгены». За последние пятнадцать-двадцать лет несколько способов было разработано для получения трансгенных растений, и настоящее изобретение, в конкретных способах осуществления, также относится к трансформированным версиям патентуемого сорта или линии. Трансформация растения включает конструирование вектора для экспрессии, который будет функционировать в растительных клетках. Такой вектор включает ДНК, включающую контролируемый ген, или функционально связанный с регуляторным элементом (например, промотором). Вектор экспрессии может содержать один или более таких функционально связанных комбинаций ген/регуляторный элемент. Вектор(ы) может быть в форме плазмиды и может быть использован самостоятельно или в комбинации с другими плазмидами, чтобы предоставить трансформированные растения подсолнечника, применяя способы трансформации, как описано ниже, для введения трансгенов в генетический материал растения(й) подсолнечника. Векторы экспрессии для трансформации подсолнечника: Маркерные Гены Векторы экспрессии включают, по меньшей мере, один генетический маркер, функционально связанный с регуляторным элементом (например, промотором), который позволяет трансформированным клеткам, содержащим маркер, быть восстановленными как негативной селекцией, т.е. ингибированием роста клеток, которые не содержат гены маркеров селекции, так и позитивной селекцией, т.е. скринингом на продукт, кодируемый генетическим маркером. Многие широко используемые гены маркеров селекции для трансформации растений хорошо известны в области трансформаций и включают, например, гены, которые кодируют ферменты, которые метаболически детоксифицируют выборочный химический агент, который может быть антибиотиком или гербицидом, или гены, которые кодируют измененную мишень, которая является не чувствительной к ингибитору. Также в уровне техники известно несколько способов позитивной селекции. Одним широко используемым геном маркеров селекции для трансформации растений является ген неомицинфосфотрансферазы II (nptII), контролируемый регуляторными сигналами растения, который отвечает за устойчивость к канамицину. Fraley et al., Proc. Natl. Acad. Sci. U.S.A., 80:4803 (1983). Другим широко используемым геном маркеров селекции является ген гигромицинфосфотрансферазы, который отвечает за устойчивость к антибиотику гигромицину. Vanden Elzen et al., Plant MoI. Biol., 5:299 (1985). Дополнительные гены маркеров селекции бактериального происхождения, которые отвечают за устойчивость к антибиотикам, включают гентамицинацетилтрансферазу, стрептомицинфосфотрансферазу и аминогликозид-3'-аденилтрансферазу, устойчивую к детерминанте блеомицина. Hayford et al., Plant Physiol. 86:1216 (1988), Jones et al., MoI. Gen. Genet, 210:86 (1987), Svab et al., Plant MoI. Biol. 14:197 (1990) HiIIe et al., Plant MoI. Biol. 7:171 (1986). Другие гены маркеров селекции отвечают за устойчивость к гербицидам, таким как глифосат, глюфосинат или броксинил. Comai et al., Nature 317:741 -744 (1985), Gordon-Kamm et al., Plant Cell 2:603-618 (1990) и Stalker et al., Science 242:419-423 (1988). Другие гены маркеров селекции для трансформации растений имеют не бактериальное происхождение. Эти гены включают, например, дигидрофолатредуктазу мыши, растительную 5-енолпирувилшикимат-3-фосфатсинтазу и растительную ацетолактатсинтазу. Eichholtz et al., Somatic Cell MoI. Genet. 13:67 (1987), Shah et al., Science 233:478 (1986), Charest et al., Plant Cell Rep. 8:643 (1990). Другой класс маркерных генов для трансформации растений требует скорее скрининга предположительно трансформированных растительных клеток, чем прямого генетического отбора трансформированных клеток, на устойчивость к токсичным веществам, таким как антибиотик. Эти гены в особенности полезны для количественного описания или визуализации пространственного паттерна экспрессии гена в специфичных тканях и часто относятся к репортерным генам, поскольку они могут быть сшиты с геном или регуляторной последовательностью гена для исследования экспрессии гена. Широко используемые гены для скрининга предположительно транформированных клеток включают Доступны способы визуализации in vivo активности GUS, которые не требуют разрушения растительной ткани. Molecular Probes publication 2908, lmagene Greenj, p. 1 -4 (1993) и Naleway et al., J. Cell Biol. 115:151a (1991). Однако эти способы визуализации in vivo активности GUS не подтвердили пригодность для выявления трансформированных клеток из-за низкой чувствительности, высокого флуоресцентного фона и ограничений, связанных с использованием генов люциферазы в качестве селектируемых маркеров. Ген, кодирующий зеленый флуоресцирующий белок (GFP), был использован в качестве маркера для экспрессии гена в прокариотических и эукариотических клетках. Chalfie et al., Science 263:802 (1994). GFP и мутированные GFP могут быть использованы в качестве скринируемых маркеров. Векторы экспрессии для трансформации подсолнечника: Промоторы Гены, включенные в векторы для экспрессии, должны быть управляемы нуклеотидной последовательностью, включающей регуляторный элемент, например промотор. Несколько типов промоторов сейчас хорошо известны в области трансформаций, как и другие регуляторные элементы, которые могут быть применены самостоятельно или к комбинации с промоторами. Как употреблено здесь, промотор включает ссылку на участок ДНК выше точки старта транскрипции и вовлеченного в распознавание и связывание РНК полимеразы и других белков для инициирования транскрипции. Промотор растения является промотором, способным инициировать транскрипцию в растительных клетках. Примеры промоторов под контролем развития включают промоторы, которые предпочтительно инициируют транскрипцию в определенных тканях, таких как листья, корни, семена, волокнах, сосудах ксилемы, трахеидах или склеренхиме. Такие промоторы относят к ткане-предпочтительным. Промоторы, которые инициируют транскрипцию только в определенных тканях, относят к ткане-специфичным. Промотор, специфичный к типу клетки, в первую очередь, управляет экспрессией в определенных типах клеток в одном или более органов, например, в сосудистых клетках в корнях или листьях. Индуцибельный промотор является промотором, который находится под контролируемыми условиями окружающей среды. Примеры условий окружающей среды, которые могут влиять на транскрипцию посредством индуцибельных промоторов, включают анаэробные условия или присутствие света. Ткане-специфичные, ткане-предпочтительные, специфичные к типу клетки и индуцибельные промоторы составляют класс неконститутивных промоторов. Конститутивный промотор является промотором, который активен при большинстве условий окружающей среды. А. Индуцибельные промоторы - индуцибельный промотор функционально связан с геном экспрессии в подсолнечнике. Необязательно, индуцибельный промотор функционально связан с нуклеотидной последовательностью, кодирующей сигнальную последовательность, которая функционально связана с геном экспрессии в подсолнечнике. С индуцибельным промотором скорость транскрипции увеличивается под влиянием индуцирующего агента. Любой индуцибельный промотор может быть применен в настоящем изобретении. См. Ward et al., Plant MoI. Biol. 22:361-366 (1993). Типичные индуцибельные промоторы включают те, которые получены из ACEI системы, которые реагируют на медь, но не ограничены ими (Mett et al., PNAS 90:4567-4571 (1993)); ген In2 из маиса, который отвечает за безопасность к бензолсульфамидному гербициду (Hershey et al., MoI. Gen Genetics 227:229-237 (1991) and Gatz et al., MoI. Gen. Genetics 243:32-38 (1994)) или репрессор Tet из Tn10 (Gatz et al., MoI. Gen. Genetics 227:229-237 (1991)). В частности, предпочтительным индуцибельным промотором является промотор, который реагирует на индуцирующий агент, на который растения обычно не реагируют. Типичным индуцибельным промотором является индуцибельный промотор из гена стероидного гормона, транскрипционная активность которого индуцируется гликокортикостероидным гормоном. Schena et al., Proc. Natl. Acad. ScL U.S.A. 88:0421 (1991). B. Конститутивные промоторы. Конститутивный промотор функционально связан с геном экспрессии в подсолнечнике, либо конститутивный промотор функционально связан с нуклеотидной последовательностью, кодирующей сигнальную последовательность, которая функционально связана с геном экспрессии в подсолнечнике. Многие различные конститутивные промоторы могут быть использованы в рассматриваемом в данный момент изобретении. Типичные конститутивные промоторы включают промоторы из растительных вирусов, но не ограничены ими, такие как промотор 35S из CaMV (Odell et al., Nature 313:810-812 (1985)) и промоторы из таких генов, как рисовый актин (McElroy et al., Plant Cell2:163-171 (1990)); убиквитин (Christensen et al., Plant MoI. Biol. 12:619-632 (1989) и Christensen et al., Plant MoI. Biol. 18:675-689 (1992)); pEMU (Last et al., Theor. Appl. Genet. 81:581 -588 (1991 )); MAS (Velten et al., EMBO J. 3:2723-2730 (1984)), H3 гистон кукурузы (Lepetit et al., MoI. Gen. Genetics 231:276-285 (1992) и Atanassova et al., Plant Journal 2 (3): 291 -300 (1992)), актин арабидопсиса (Last and Gray, Plant MoI. Biol. 12:655-666 (1989)) и промотор пластоцианина гороха (McCabe et al., Theor. Appl. Genet. 99:587- 592 (1999)). ALS промотор, фрагмент 5' XbaI/NcoI, к структурному гену ALS3 Brassica napus (или нуклеотидной последовательности, схожей с указанным XbaI/NcoI фрагментом), представляет собой особенно важный конститутивный промотор. Смотри заявку PCT WO 96/30530. С. Ткане-специфичные или ткане-предпочтительные промоторы. Ткане-специфичный промотор функционально связан с геном экспрессии в подсолнечнике. Необязательно, ткане-специфичный промотор функционально связан с нуклеотидной последовательностью, кодирующей сигнальную последовательность, которая функционально связана с геном экспрессии в подсолнечнике. Растения, трансформированные интересующим геном, функционально связанным с ткане-специфичным промотором, продуцируют белковый продукт трансгена эксклюзивно или предпочтительно в специфической ткани. Любой ткане-специфичный или ткане-предпочтительный промотор может быть применен в настоящем изобретении. Типичные ткане-специфичные или ткане-предпочтительные промоторы включают, но не ограничены ими, корне-предпочтительный промотор, такой как из гена фазеолина (Murai et al., Science 23:476-482 (1983) и Sengupta-Gopalan et al., Proc. Natl. Acad. Sci. U.S.A. 82:3320-3324 (1985)); листо-специфический и свето-индуцированный промотор, такой как из капусты или цветной капусты (Simpson et al., EMBO J. 4(11):2723-2729 (1985) и Timko et al., Nature 318:579-582 (1985)); пыльнико-специфический промотор, такой как из LAT52 (Twell et al., MoI. Gen. Genetics 217:240-245 (1989)); a пыльце-специфический промотор, такой как из Zm13 (Guerrero et al., MoI. Gen. Genetics 244:161 -168 (1993)) или a микроспоро-предпочтительный промотор, такой как из apg (Twell et al., Sex. Plant Reprod. 6:217-224 (1993)). Транспорт белка, продуцированного трансгенами в субклеточные компартменты, такие как хлоропласты, вакуоли, пероксисомы, глиоксисомы, клеточная стенка или митохондрия, или для секреции в апопласт, выполняется посредством функционального связывания нуклеотидной последовательности, кодирующей последовательность сигнала к 5' и/или 3' участку гена, кодирующего интересующий белок. Нацеливающие последовательности на 5' и/или 3' конце структурного гена могут определять, во время синтеза белка и процессирования, где закодированный белок в итоге будет компартментализирован. Наличие сигнальной последовательности направляет полипептид как к внутриклеточной органелле или субклеточному компартменту, так и для секреции в апопласт. В уровне техники известно много сигнальных последовательностей. См., например, Becker et al., Plant MoI. Biol. 20:49 (1992), Close, P.S., Master's Thesis, Iowa State University (1993), Knox, C, et al., AStructure and Organization of Two Divergent Alpha-Amylase Genes from Barley®, Plant MoI. Biol. 9:3- 17 (1987), Lerner et al., Plant Physiol. 91:124-129 (1989), Fontes et al., Plant Cell 3:483-496 (1991 ), Matsuoka et al., Proc. Natl. Acad. ScL 88:834 (1991 ), Gould et al., J. Cell. Biol. 108:1657 (1989), Creissen et al., Plant J. 2:129 (1991 ), Kalderon, et al., A short amino acid sequence able to specify nuclear location. Cell 39:499-509 (1984), Steifel, et al., Expression of a maize cell wall hydroxyproline-rich glycoprotein gene in early leaf and root vascular differentiation, Plant Cell 2:785-793 (1990). Гены чужеродных белков и агрономические гены С трансгенными растениями по настоящему изобретению чужеродный белок может быть получен в коммерческих количествах. Таким образом, методы для селекции и разведения трансформированных растений, хорошо известные в уровне техники, дают множество трансгенных растений, урожай которых собирается традиционным путем, и чужеродный белок далее может быть экстрагирован из интересующей ткани или из всей биомассы. Экстракция белка из растительной биомассы может быть выполнена известными способами, которые обсуждаются, например, Heney and Orr, Anal. Biochem. 114:92-6 (1981). Согласно предпочтительному способу осуществления, трансгенным растением, предусмотренным для коммерческого получения чужеродного белка, является растение подсолнечника. В другом предпочтительном способе осуществления интересующей биомассой являются семена. Для относительного малого количества трансгенных растений, которые показывают более высокие уровни экспрессии, может быть создана генетическая карта, в первую очередь, по традиционным RFLP, PCR и SSR анализам, которые идентифицируют приблизительное положение на хромосоме интегрированной молекулы ДНК. Примеры методик в этом направлении, см. Glick and Thompson, Methods in Plant Molecular Biology and Biotechnology CRC Press, Boca Raton 269:284 (1993). Информация карты, касающаяся положения на хромосоме, является полезной для защиты патентуемого объекта - трансгенного растения. Если предпринимается неправомерное разведение и скрещивание ведется с другой зародышевой плазмой, карта района интеграции может сравниваться со схожими картами для предполагаемых растений, для определения, имеют ли последние общее происхождение с испытуемым растением. Сравнение карт вовлекло бы скрещивания, RFLP, PCR и SSR и секвенирование - все из которых являются традиционными методами. Подобным образом, посредством настоящего изобретения, агрономические гены могут быть экспрессированы в трансформированных растениях. Более конкретно, растения могут быть созданы методами генной инженерии для экспрессирования разных фенотипов, представляющих агрономический интерес. Типичные гены, вовлеченные при этом, включают гены, упорядоченные ниже по категориям, но не ограничиваются ими: 1. Гены, отвечающие за устойчивость к вредителям или болезням и которые кодируют: А. Гены устойчивости к заболеваниям растений. Защитные свойства растения часто активируются специфическим взаимодействием между продуктом гена устойчивости к заболеваниям (R) растения и продуктом соответствующего гена авирулентности (Avr) в патогене. Сорт растения может быть трансформирован клонированными генами устойчивости растения, созданными методами генной инженерии, устойчивыми к специфичным патогенным штаммам. См., например, Jones et al., Science 266:789 (1994) (Клонирование гена томата Cf-9 для устойчивости к Cladosporium fulvum); Martin et al., Science 262: 1432 (1993) (Ген томата Pto для устойчивости к Pseudomonas syringae pv. Томат кодирует протеинкиназу); Mindrinos et al., Cell 78:1089 (1994) (Ген арабидопсиса RSP2 для устойчивости к Pseudomonas syringae). B. Ген, отвечающий за устойчивость к вредителям, таким как нематоды цисты соевых бобов. См., например, заявку PCT WO 96/30517; заявку PCT WO 93/19181. C. Белок Bacillus thuringiensis, его производное или синтетический полипептид, моделирующий его. См., например, Geiser et al., Gene 48: 109 (1986), который описывает клонирование и нуклеотидную последовательность гена Bt D. Лектин. См., например, Van Damme et al., Plant Molec. Biol. 24:25 (1994), который описывает нуклеотидные последовательности нескольких генов маннозо-связывающего лектина Clivia miniata. E. Витамин-связывающий белок, такой как авидин. Смотри заявку PCT US93/06487. В заявке сообщается об использовании авидина и гомологов авидина в качестве ларвицидов против насекомых вредителей. F. Ингибитор фермента, например, ингибитор протеазы или протеиназы, или ингибитор амилазы. См., например, Abe et al., J. Biol. Chem. 262:16793 (1987) (последовательность нуклеотидов ингибитора цистеиновой протеиназы риса), Huub et al., Plant Molec. Biol. 21:985 (1993) (последовательность нуклеотидов кДНК, кодирующей ингибитор I табачной протеиназы), Sumitani et al., Biosci. Biotech. Biochem. 57:1243 (1993) (последовательность нуклеотидов ингибитора G. Насекомо-специфичный гормон или феромон, такой как экдистероид и ювенильный гормон, их варианты, миметики, или их антагонист или агонист. См., например, описание в Hammock et al., Nature 344:458 (1990), экспрессии клонированной эстеразы ювенильного гормона бакуловируса, инактиватора ювенильного гормона. H. Насекомо-специфичный белок или нейропептид, который при экспрессии нарушает физиологию подверженного влиянию вредителя. Например, смотри описания Regan, J. Biol. Chem. 269:9 (1994) (выходом экспрессионного клонирования является ДНК, кодирующая диуретический рецептор гормона насекомого), и Pratt et al., Biochem. Biophys. Res. Comm. 163:1243 (1989) (аллостатин идентифицирован в Diploptera puntata). Смотри также Патент США No. 5266317, выданный Tomalski et al., который описывает гены, кодирующие насекомо-специфичные паралитические нейротоксины. I. Насекомо-специфичный яд, вырабатываемый в природе змеями, осами и т.д. Например, смотри Pang et al., Gene 116:165 (1992) для описания гетерологичной экспрессии в растениях гена, кодирующего насекомотоксичный белок скорпиона. J. Фермент, отвечающий за гипераккумуляцию монотерпена, сесквитерпена, стероида, гидроксамовой кислоты, производных фенилпропаноида или другой небелковой молекулы с инсектицидным действием. K. Фермент, вовлеченный в модификацию, включающий посттрансляционную модификацию биологически активных молекул; например гликолитический фермент, протеолитический фермент, липолитический фермент, нуклеаза, циклаза, трансаминаза, эстераза, гидролаза, фосфатаза, киназа, фосфорилаза, полимераза, эластаза, хитиназа и глюканаза, натуральные либо синтетические. Смотри заявку PCT WO 93/02197 на имя Scott et al., который описывает нуклеотидную последовательность гена каллазы. Молекулы ДНК, которые содержат последовательности, кодирующие хитиназу, могут быть получены, например, в ATCC под номерами доступа 39637 и 67152. См. также Kramer et al., Insect Biochem. Molec. Biol. 23:691 (1993), который показывает нуклеотидную последовательность кДНК, кодирующей хитиназу табачной анкилостомозы, и Kawalleck et al., Plant Molec. Biol. 21:673 (1993), который приводит нуклеотидную последовательность гена ubi4-2 полиубиквитина петрушки. L. Молекула, которая стимулирует сигнальную трансдукцию. Например, смотри описание нуклеотидной последовательности для клонов кальмодулин кДНК золотистой фасоли Botella et al., Plant Molec. Biol. 24:757 (1994)и Griess et al., Plant Physiol. 104:1467 (1994), который представил нуклеотидную последовательность слона кальмодулина кДНК кукурузы. M. Гидрофобный влияющий белок. Смотри заявку PCT WO 95/16776 (описание белковых производных тахиплезина, которые ингибируют грибные патогенные растения) и заявку PCT WO 95/18855 (показывает синтетические антибактериальные белки, отвечающие за устойчивость к инфекциям). N. Мембранная пермеаза, образователь или блoкатор каналов. Например, смотри описание гетерологичной экспрессии цекропина- O. Вирусно-инвазивный белок или получаемый из него сложный яд. Например, аккумуляция белков оболочки вирусов в клетках трансформированных растений придает устойчивость к вирусной инфекции и/или развитию заболевания под влиянием вируса, из которого вырабатывается ген белка оболочки так же, как из связанных вирусов. См. Beachy et al., Ann. rev. Phytopathol. 28:451 (1990). Устойчивость, опосредованная белком оболочки, придаваемая трансформированным растениям устойчивость против вируса мозаики люцерны, вируса мозаики огурца, вируса табачной полоски, картофельного вируса X, картофельного вируса Y, вируса протравы табака, вируса треска табака и вируса табачной мозаики Id. P. Насекомо-специфичное антитело или иммунотоксин, получаемый из него. Так, антитело, нацеленное на критическую метаболическую функцию внутри насекомого, могло бы инактивировать фермент, находящийся под влиянием, убивая насекомое. См. Taylor et al., Abstract #497, Seventh Int'l Symposium on Molecular Plant- Microbe Interactions (Edinburgh, Scotland) (1994) (инактивация фермента в трансгенном табаке через получение одноцепочечных фрагментов антитела). Q. Вирус-специфичное антитело. Смотри, например, Tavladoraki et al., Nature 366:469 (1993), который показывает, что трансгенные растения, экспрессирующие гены рекомбинантного антитела, защищены от атаки вируса. R. Останавливающий развитие белок, продуцируемый в природе патогеном или паразитом. Так, грибные эндо S. Останавливающий развитие белок, продуцируемый в природе растением. Например, Logemann et al., Bio/Technology 10:305 (1992), показали, что трансгенные растения, экспрессирующие ген инактивации рибосом ячменя, обладают повышенной устойчивостью к грибным заболеваниям. 2. Гены, ответственные за устойчивость к гербициду: A. Гербицид, который ингибирует точку роста или меристему, такой как имидазолинон или сульфонилмочевина. Типичные гены этой категории кодируют мутированный ALS и фермент AHAS, как описано, например, Lee et al., EMBO J. 7:1241 (1988), и Miki et al., Theor. Appl. Genet. 80:449 (1990), соответственно. B. Глифосат (устойчивость, ослабленная генами мутированной 5-енолпирувил-3-фосфошикиматсинтазы (EPSP) и ароA, соответственно) и другие фосфоно-соединения, такие как глюфосинат (фосфинотрицинацетилтрансфераза, (PAT) и фосфинотрицинацетилтрансфераза Streptomyces hygroscopicus bar гены), и пиридинокси- или фенокси-пропионовые кислоты и циклогексоны (гены, кодирующие ингибитор ACCase). См., например, Патент США No. 4940835, выданный Shah, et al., который описывает нуклеотидную последовательность формы EPSP, которая может отвечать за устойчивость к глифосату. Молекула ДНК, кодирующая мутированный ген ароА, может быть получена в ATCC с номером доступа 39256, и нуклеотидная последовательность мутированного гена описывается в Патенте США No. 4769061, выданном Comai. Европейская патентная заявка No. 0333033, выданная Kumada et al., и Патент США No. 4975374, выданный Goodman et al., описывают нуклеотидные последовательности генов глутаминсинтетазы, которые отвечают за устойчивость к гербицидам, таким как L-фосфинотрицин. Нуклеотидная последовательность гена фосфинотрицинацетилтрансферазы приводится в Европейской Патентной заявке No. 0242246, выданной Leemans et al., DeGreef et al., Bio/Technology 7:61 (1989), описывает получение трансгенных растений, которые экспрессируют химерные bar гены, кодирующие фосфинотрицинацетилтрансферазную активность. Типичными генами, отвечающими за устойчивость к феноксипропионовым кислотам и циклогексонам, таким как сетоксидим и галоксифоп, являются гены Acc1-S1, Acc1-S2 и Acc1-S3, описанные Marshall et al., Theor. Appl. Genet. 83:435 (1992). C. Гербицид, ингибирующий фотосинтез, такой как триазин (psbA и gs+ гены) или бензонитрил (ген нитрилазы). Przibila et al., Plant Cell 3: 169 (1991), описывают трансформацию Chlamydomonas с плазмидами, кодирующими мутированные гены psbA. Нуклеотидные последовательности для генов нитрилаз описаны в Патенте США No. 4810648, выданном Stalker, и молекулы ДНК, содержащие эти гены, доступны в ATCC под номерами доступа 53435, 67441 и 67442. Клонирование и экспрессия ДНК, кодирующей a глутатион S-трансфераза, описаны Hayes et al., Biochem. J. 285:173 (1992). 3. Гены, которые отвечают за или делают вклад в признаки добавления стоимости, такие как: A. Модифицированный метаболизм жирных кислот, например, путем трансформирования растения с антисмысловым геном стеароил-ACP дезатуразы для увеличения содержания стеариновой кислоты в растении. См. Knultzon et al., Proc. Natl. Acad. Sci. U.S.A. 89:2624 (1992). B. Сниженное содержание фитата. 1. Введение гена, кодирующего фитазу, увеличило бы деградацию фитата, добавляя больше свободного фосфата в трансформированное растение. Например, смотри Van Hartingsveldt et al., Gene 127:87 (1993) для описания нуклеотидной последовательности гена фитазы Aspergillus niger. 2. Может быть введен ген, который уменьшает содержание фитата. В кукурузе, например, это может быть выполнено клонированием и дальнейшим введением ДНК, ассоциированной с одиночной аллелью, которая отвечает за мутанты кукурузы, характеризующиеся низкими содержаниями фитиновой кислоты. Смотри Raboy et al., Maydica 35:383 (1990). C. Модифицированный углеводный состав растений, осуществляемый, например, трансформированным геном, кодирующим фермент, который изменяет ветвистую сеть крахмала. См. Shiroza et al., J. Bacteol. 170:810 (1988) (нуклеотидная последовательность гена фруктозилтрансферазы Streptococcusmutants), Steinmetz et al., MoI. Gen. Genet. 20:220 (1985) (нуклеотидная последовательность гена левансукразы Bacillus subtilis), Pen et al., Bio/Technology Способы трансформации подсолнечника Разработаны многочисленные способы трансформации растений, включая биологические и физические, протоколы трансформации растений. См., например, Miki et al., "Procedures for Introducing Foreign DNA into Plants" In Methods in Plant Molecular Biology and Biotechnology, Glick B. R. и Thompson, J. E. Eds. (CRC Press, Inc., Boca Raton, 1993) pp. 67-88. В дополнение, векторы для экспрессии и in vitro методы культивирования для растительной клетки или трансформации ткани и регенерации растений являются доступными. См., например, Gruber et al., "Vectors for Plant Transformation" in Methods in Plant Molecular Biology and Biotechnology, Glick B. R. и Thompson, J. E. Eds. (CRC Press, Inc., Boca Raton, 1993) pp. 89-119. A. Agrobacterium-опосредованная трансформация - один из способов введения вектора экспрессии в растения, основанный на естественной системе трансформации grobacterium. См., например, Horsch et al., Science 227:1229 (1985). A. tumefaciens и A. rhizogenes являются патогенными почвенными бактериями растений, которые генетически трансформируют растительные клетки. Ti и Ri плазмиды для A. tumefaciens и A. rhizogenes, соответственно, несут гены, отвечающие за генетическую трансформацию растений. См., например, Kado, C. I., Crit. Rev. Plant Sci. 10:1 (1991). Описания системы векторов для Agrobacterium и способов для Agrobacterium-опосредованного переноса гена приводятся Gruber et al., supra, Miki et al., supra, и Moloney et al., Plant Cell Reports 8:238 (1989). Смотри также Патент США No. 5563055 (Townsend and Thomas), выданный 8 Октября, 1996. B. Прямой перенос гена. Несколько способов трансформации растения, вместе относящихся к прямому переносу гена, были разработаны в качестве альтернативы трансформации, опосредованной Agrobacterium. Широко применяемым способом трансформации растений является микроинжекторно-опосредованная трансформация, где ДНК наносится на поверхность микроинжекторного устройства размером от 1 до 4 мкм. Вектор экспрессии вводится в растительные ткани при помощи биолистического аппарата, который ускоряет микроинжекторы до скорости 300-600 м/с, которая является достаточной для проникновения через растительные клеточные стенки и мембраны. Sanford et al., Part. Sci. Technol. 5:27 (1987), Sanford, J.C., Trends Biotech. 6:299 (1988), Klein et al., Bio/Technology 6:559-563 (1988), Sanford, J.C., Physiol Plant 7:206 (1990), Klein et al., Biotechnology 10:268 (1992). Смотри также Патент США No.5015580 (Christou, et al.), выданный 14 мая, 1991; Патент США No. 5322783 (Tomes, et al.), выданный 21 июня, 1994. Другим способом физического внесения ДНК в растения является разрушение ультразвуком клеток-мишеней. Zhang et al., Bio/Technology 9:996 (1991). Альтернативно, было использовано слияние липосом или сферопластов для введения векторов экспрессии в растения. Deshayes et al., EMBO J., 4:2731 (1985), Christou et al., Proc Natl. Acad. Sci. U.S.A. 84:3962 (1987). Прямое поглощение ДНК протопластами при использовании осаждения CaCl2, поливинилового спирта или поли-L-омитина также были представлены. Hain et al., MoI. Gen. Genet. 199:161 (1985) и Draper et al., Plant Cell Physiol. 23:451 (1982). Электропорация протопластов и целых клеток, и тканей также была описана. Donn et al., In Abstracts of VIIth International Congress on Plant Cell and Tissue Culture IAPTC, A2-38, с. 53 (1990); D'Halluin et al., Plant Cell 4:1495-1505 (1992) and Spencer et al., Plant Mol. Biol. 24:51 - 61 (1994). После трансформации ткани-мишени подсолнечника экспрессия вышеописанных генов маркеров селекции учитывает предпочтительный отбор трансформированных клеток, тканей и/или растений, используя способы регенерации и селекции, хорошо известные в данной области техники. Вышеуказанные способы для трансформации обычно применяются для получения трансгенных сортов. Трансгенный сорт далее может быть скрещен с другим (нетрансформированным или трансформированным) сортом, чтобы получить новый трансгенный сорт. Альтернативно, генетический признак, который был встроен в конкретную линию подсолнечника, используя предыдущие способы трансформации, может быть перенесен в другую линию, используя традиционные способы обратного скрещивания, которые хорошо известны в области селекции растений. Например, подход обратного скрещивания мог бы быть применен для переноса встроенного признака из общественного, неэлитного сорта в элитный сорт, или из сорта, содержащего чужеродный ген в своем геноме, в сорт или сорта, которые не содержат тот ген. Как употреблено здесь, "скрещивание" может относиться к простому X и Y скрещиванию или процессу обратного скрещивания, зависящему от контекста Тканевая культура подсолнечника. Дальнейшее получение сортов OI1601A, OI2653R и OI1601B или гибрида E83329 может происходить самоопылением или тканевой культурой и регенерацией. Известна тканевая культура различных тканей подсолнечника и регенерация растений из них. Например, разведение сорта подсолнечника тканевой культурой описано в любом из следующих, но не ограничиваясь перечисленным: Shin et al., In Vitro Cellular and Development Biology - Plant, 36:273-278 (2000); Hildebrandt и Riker, Amer. J. Bot, 34:421 -427 (1947); Rogers et al., In Vitro, 6:463-7 (1974); Fambrini et al., Ann. Bot, 92:145-152 (2003). Когда термин «растение подсолнечника» употребляется в контексте настоящего изобретения, это также включает любые конверсии одиночного гена этого сорта. Термин «растение с одиночным конвертированным геном», как употреблено здесь, относится к тем растениям подсолнечника, которые получены методами селекции растений, называемыми обратным скрещиванием, где все существенно желаемые морфологические и физиологические характеристики сорта получены в дополнение к одиночному гену, перенесенному в сорт посредством метода обратного скрещивания. Методы обратного скрещивания могут быть использованы в настоящем изобретении для улучшения или введения характеристики в сорт. Термин «Обратное скрещивание», как употреблено здесь, относится к многократному возвратному скрещиванию гибридного потомства с рекуррентным родителем, т.е обратное скрещивание 1, 2, 3, 4, 5, 6, 7, 8 или более раз с рекуррентным родителем. Родительское растение подсолнечника, которое вносит в ген желаемую характеристику, называется "нерекуррентным Выбор подходящего рекуррентного родителя является важным шагом для успешной процедуры обратного скрещивания. Целью протокола обратного скрещивания является изменение или замена одиночного признака или характеристики в исходном сорте. Для выполнения этого одиночный ген рекуррентного сорта модифицируется или заменяется желаемым геном из нерекуррентного родителя, при этом оставляя существенно весь остальной желаемый генетический, и потому желаемый физиологический и морфологический, состав исходного сорта. Выбор конкретного нерекуррентного родителя будет зависеть от цели обратного скрещивания. Одной из главных целей является добавление некоторого коммерчески желаемого, агрономически важного признака растения. Точный протокол обратного скрещивания будет зависеть от характеристики или признака изменяемого для определения подходящего тестирующего протокола. Хотя методы обратного скрещивания упрощаются, когда переносимая характеристика представляет собой доминантную аллель, рецессивная аллель также может быть перенесена. В этом случае может быть необходимым введение теста потомства для того, чтобы определить, была ли желаемая характеристика успешно перенесена. Идентифицировано много признаков одиночного гена, которые обычно не выбираются при разработке нового сорта, но которые могут быть улучшены методами обратного скрещивания. Признаки одиночного гена могут быть или могут не быть трансгенными, примеры этих признаков включают мужскую стерильность, восковость крахмала, устойчивость к гербицидам, устойчивость к бактериальным, грибным или вирусным заболеваниям, устойчивость к насекомым, мужскую оплодотворяющую способность, повышенное качество питательности, промышленную применимость, устойчивость урожая и повышенную урожайность, но не ограничены ими. Эти гены в основном наследуются через ядро. Некоторые из этих признаков одиночного гена описаны в Патентах США No.No.5959185, 5973234 и 5977445. Дальнейшее воспроизводство сорта может происходить тканевой культурой или регенерацией. Тканевые культуры различных тканей подсолнечника и регенерированные из них растения хорошо известны и широко публикуются. Например, ссылкой может быть Mayor, et al., Plant Cell, Tissue and Organ Culture, 72:99-103 (2003) и Baker et al., Plant Cell, Tissue and Organ Culture, 58:39-49 (1999). Таким образом, другим аспектом этого изобретения является предоставление клеток, которые после выращивания и дифференциации воспроизводят растения подсолнечника, обладающие физиологическими и морфологическими характеристиками сортов подсолнечника OI1601A, OI2653R или OI1601B, или гибрида подсолнечника E83329. Как употреблено здесь, термин тканевая культура обозначает смесь, включающую изолированные клетки одного и того же или разного вида, или скопление таких клеток, организованных в части растения. Типичными видами тканевых культур являются протопласты, каллус, растительные куски и клетки растения, которые могут генерировать цельную тканевую культуру в растениях или частях растений, такую как эмбрионы, пыльца, цветки, семена, стручки, листья, стебли, корни, кончики корней, пыльники и подобные. Средства подготовки и поддержания растительной тканевой культуры хорошо известны в данной области техники. В качестве примера, тканевая культура, включающая органы, была применена для получения регенерированных растений. Патенты США No.No.5959185; 5973234 и 5977445 описывают конкретные способы, описания которых включены ссылкой. Это изобретение также направлено на способы получения растения подсолнечника путем скрещивания первого родительского растения подсолнечника со вторым родительским растением подсолнечника, где первое или второе родительское растение подсолнечника является сортом растения подсолнечника OI1601A, OI2653R или OI1601B, или гибридом E83329. Дополнительно, оба - первое и второе, родительские растения подсолнечника могут происходить из сорта подсолнечника OI1601A, OI2653R или OI1601B или гибрида E83329. Таким образом, любые такие способы с использованием сорта подсолнечника OI1601A, OI2653R или OI1601B или гибрида подсолнечника E83329 являются частью этого изобретения: селфинг, обратные скрещивания, получение гибридов, скрещивание популяций и подобные. Все растения, полученные при использовании сорта подсолнечника OI1601A, OI2653R или OI1601B или гибрида подсолнечника E83329, в качестве родителя, находятся в объеме этого изобретения, включая полученные из сортов, полученных из сорта подсолнечника OI1601A, OI2653R или OI1601B или гибрида подсолнечника E83329. Преимущественно, сорт подсолнечника мог бы быть применен при скрещиваниях с другими, различными растениями подсолнечника для получения первого поколения (F1) гибридных семян подсолнечника и растений с превосходными характеристиками. Сорт данного изобретения также может быть применен для трансформации, где экзогенные гены вводятся и экспрессируются сортом данного изобретения. Генетические варианты, созданные как традиционной селекцией с применением сортов OI1601A, OI2653R или OI1601B или гибрида E83329, так и трансформацией сортов OI1601A, Ol2653R или OI1601B или гибрида E83329 любым из протоколов, известных специалистам в данной области техники, предполагаются находящимися в пределах объема настоящего изобретения. Депозиты сортов подсолнечника OI1601A, OI2653R или OI1601B или гибрида подсолнечника E83329 сохраняются в Dow Agrosciences at Agrigenetics, Inc. d/b/a Mycogen Seeds, Highway 75 North, Breckenridge, Minnesota 56520. Доступ к этим депозитам будет открыт при рассмотрении этой заявки личностям, определенным быть уполномоченными на это, Комиссаром по Патентам и торговым Маркам, под 37 CFR 1.14 и 35 USC 122. После принятия любых пунктов формулы изобретения в этой заявке все ограничения по доступности этих сортов для общественности будут безвозвратно сняты путем обеспечения доступа к депонированным, по меньшей мере, 2500 семянам каждого из перeчисленных выше сортов в Американской Коллекции типовых Культур - Манассас, Вирджиния. В то время как несколько типовых аспектов и способов осуществления описывались выше, специалисты в данной области техники распознают их определенные модификации, пермутации, дополнения и подкомбинации. Поэтому предполагается, что следующие прилагаемые пункты формулы изобретения и пункты формулы изобретения, далее внесенные на рассмотрение, толкуются включающими все подобные модификации, изменения, дополнения и подкомбинации как находящиеся в пределах их истинной сущности и объема. Формула изобретения1. Способ получения гибридного семени подсолнечника, где способ включает скрещивание инбредного растения подсолнечника, который характеризуется повышенным содержанием олеиновой кислоты и устойчивостью к имидазолинону, выбранного из сортов OI601 A, OI2653R, OI601В или Е83329, продуцирующего семя подсолнечника, депонированное в АТСС под депозитным номером РТА-9470, РТА-9472, РТА-9471 или РТА-9473 соответственно с иным инбредным растением подсолнечника и сбор полученного гибридного семени подсолнечника. 2. Способ по п.1, где полученное гибридное семя подсолнечника имеет содержание олеиновой кислоты приблизительно более чем 80% по массе. 3. Способ по п.1, дополнительно включающий дробление полученного гибридного семени подсолнечника для получения масла, имеющего содержание олеиновой кислоты приблизительно более чем 80% по массе. Популярные патенты: 2395497 Способ стимулирования роста подсолнечника регулятором роста ... формулы 1-2: 1 R = фтор;2 R = нитро,путем предпосевной обработки семян. MM4A Досрочное прекращение действия патента из-за неуплаты в установленный срок пошлины за поддержание патента в силе Дата прекращения действия патента: 15.10.2010 Дата публикации: ... 2296457 Устройство для магнитно-импульсной обработки растений ... первый короткий импульс второй пачки импульсов N2э (фиг.2, поз.47), поступивший на управляющий вход второго ключа 13, открывает его. При этом происходит очередной разряд первого накопительного конденсатора 8 через обмотку второй катушки L2 индуктора 20 также за время к (фиг.2, поз.50). В результате во второй катушке L2 индуктора 20 возникает очередной импульс магнитной индукции (фиг.2, поз.51), воздействуя на размещенные в нем растения. Далее процесс идет аналогично и окончательно завершается с приходом заднего фронта строб-импульса длительности э с выхода цифрового таймера 30 (фиг 2, поз.42), который своим задним фронтом закрывает третий ключ 17 и прерывает подачу напряжения ... 2121263 Способ лесоводственной оценки технологического комплекса машин ... технике и технологии лесозаготовок предъявляются лесоводственные требования по условиям проведения работ в зависимости от видов рубок, состояния лесной среды до начала их проведения, а также к ее состоянию после проведения рубок [3]. Указывается, что положения и нормативы, приведенные в данном документе, должны использоваться для оценки пригодности (возможности и целесообразности использования) машин и технологией на лесосечных работах при различных способах рубок главного пользования в определенных природных (типологических условиях и с учетом сезонов года. Отмечается, что при выборе следует отдавать предпочтение тем лесозаготовительным машинам и технологиям разработки лесосек, ... 2420060 Способ генетической трансформации растений селекционно-ценных образцов клевера лугового ... способ генетической трансформации растений селекционно-ценных образцов клевера лугового позволяет получать растения-регенеранты со встроенными генами - маркерным (канамицинустойчивость) и целевым (ген синтеза дефензина амаранта, повышающим устойчивость растений к корневым гнилям).Кроме того, использование при генетической трансформации способа прямой регенерации растений, исключающего возникновение сомаклональной изменчивости, позволяет 1) сохранять селекционно-ценные признаки исходного образца (кислотоустойчивость), 2) поддерживать в культуре in vitro более 50 пассажей трансгенные морфогенные культуры с высокой регенерационной способностью, 3) получать неограниченное ... 2235464 Гербицидно-действующее средство ... соотношение гербицид формулы (II):дальнейший гербицид составляет 1:2,4.6. Способ борьбы с сорняками в культурах полезных растений путем обработки сорняков, культурных растений, семян последних или поверхности почвы, где растут растения, гербицидно-действующим веществом в гербицидно-действующем количестве, отличающийся тем, что в качестве гербицидно-действующего вещества используют средство согласно одному из пп.1-5.7. Способ по п.6, отличающийся тем, что растения производятся из группы, включающей кукурузу и пшеницу. TZ4A - Поправки к описаниям изобретений Часть описания, где обнаружена ошибка: Текст опис., Кол.7, строка 22 Напечатано: “Chemlsche Technologle” ... |
Еще из этого раздела: 2248352 Замещенные бензоилциклогександионы, гербицидное средство на их основе, исходное соединение 2140738 Производные n-арилгидразина, способ их получения, способ подавления насекомых и композиция для подавления насекомых 2280351 Установка для скашивания сорной растительной массы с берм и откосов канала 2054872 Гербицидная композиция и способ борьбы с сорняками 2404898 Устройство на воздушной подушке для разбрасывателей органоминеральных удобрений 2076603 Способ повышения урожайности сельскохозяйственных культур 2076583 Способ выращивания растений в теплице и устройство для его осуществления 2490849 Способ переработки безподстилочного помета птиц клеточного содержания и навоза свиней в топливные брикеты 2427121 Почвообрабатывающий агрегат 2279799 Балансир рыболовный |
Изобретения в сельском хозяйстве
Обработка почвы в сельском и лесном хозяйствах
Посадка, посев, удобрение
Уборка урожая, жатва
Обработка и хранение продуктов полеводства и садоводства
Садоводство, разведение овощей, цветов, риса, фруктов, винограда, лесное хозяйство
Новые виды растений или способы их выращивания
Производство молочных продуктов
Животноводство, разведение и содержание птицы, рыбы, насекомых, рыбоводство, рыболовство
Поимка, отлов или отпугивание животных
Консервирование туш животных, или растений или их частей
Биоцидная, репеллентная, аттрактантная или регулирующая рост растений активность химических соединений или препаратов
Хлебопекарные печи, машины и прочее оборудование для хлебопечения
Машины или оборудование для приготовления или обработки теста
Обработка муки или теста для выпечки, способы выпечки, мучные изделия
|
|
||
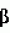 -глюкуронидазу (GUS),
-глюкуронидазу (GUS), 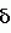 -эндотоксина. Более того, молекулы ДНК, кодирующие гены
-эндотоксина. Более того, молекулы ДНК, кодирующие гены 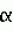 -амилазы Streptomyces nitrosporeus) и Патент США No. 5494813 (Hepher and Atkinson, выданный 27 Февраля, 1996).
-амилазы Streptomyces nitrosporeus) и Патент США No. 5494813 (Hepher and Atkinson, выданный 27 Февраля, 1996).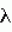 0:292 (1992) (получение трансгенных растений, которые экспрессируют
0:292 (1992) (получение трансгенных растений, которые экспрессируют  или "родителем-донором". Эта терминология относится к тому факту, что нерекуррентный родитель употребляется один раз в протоколе обратного скрещивания и поэтому не повторяется. Родительское растение подсолнечника, в которое переносится ген или гены из нерекуррентного родителя, известно как рекуррентный родитель, поскольку оно используется для нескольких циклов в протоколе обратного скрещивания (Poehlman & Sleper, 1994; Fehr, 1987). В типичном протоколе обратного скрещивания исходный интересующий сорт (рекуррентный родитель) скрещивается со вторым сортом (нерекуррентным родителем), который несет единственный интересующий ген для переноса. Полученное потомство от этого скрещивания затем снова скрещивается с рекуррентным родителем, и процесс повторяется, пока не будет получено растение подсолнечника, в котором существенно все желаемые морфологические и физиологические характеристики рекуррентного родителя получают в конвертированном растении в дополнение к одиночному гену из нерекуррентного родителя, как определено при 5-процентном уровне значимости, при выращивании в одинаковых условиях окружающей среды.
или "родителем-донором". Эта терминология относится к тому факту, что нерекуррентный родитель употребляется один раз в протоколе обратного скрещивания и поэтому не повторяется. Родительское растение подсолнечника, в которое переносится ген или гены из нерекуррентного родителя, известно как рекуррентный родитель, поскольку оно используется для нескольких циклов в протоколе обратного скрещивания (Poehlman & Sleper, 1994; Fehr, 1987). В типичном протоколе обратного скрещивания исходный интересующий сорт (рекуррентный родитель) скрещивается со вторым сортом (нерекуррентным родителем), который несет единственный интересующий ген для переноса. Полученное потомство от этого скрещивания затем снова скрещивается с рекуррентным родителем, и процесс повторяется, пока не будет получено растение подсолнечника, в котором существенно все желаемые морфологические и физиологические характеристики рекуррентного родителя получают в конвертированном растении в дополнение к одиночному гену из нерекуррентного родителя, как определено при 5-процентном уровне значимости, при выращивании в одинаковых условиях окружающей среды.