Фрагмент гибридного токсина bacillus thuringiensis, обладающий инсектицидной активностью, кодирующая его рекомбинантная днк, способ борьбы с насекомымиПатент на изобретение №: 2210593 Автор: БОШ Хендрик Ян (NL), СТИКЕМА Виллем Йоханнес (NL) Патентообладатель: НОВАРТИС АГ (CH) Дата публикации: 20 Августа, 2003 Начало действия патента: 1 Сентября, 1994 Адрес для переписки: 101000, Москва, М.Златоустинский пер., 10, кв.15, "ЕВРОМАРКПАТ", пат.пов. И.А.Веселицкой, рег. № 0011 Изображения    Изобретение относится к области биотехнологии, а именно к фрагментам гибридных токсинов, полученных из кристаллических белков Bacillus thuringiensis, обладающих инсектицидной активностью. Данные фрагменты гибридных токсинов получены посредством единичного случая рекомбинации и содержат на своем С-терминальном конце домен III CryC или CryA белка и на N-терминальном конце домен II Cry белка. Изобретение также касается рекомбинатной ДНК, кодирующей указанные фрагменты гибридных токсинов, способа борьбы с насекомыми путем их обработки фрагментами гибридных токсинов или композициями, содержащими эти токсины. Изобретение позволяет расширить арсенал эффективных средств, используемых для борьбы с насекомыми. 3 с. и 9 з.п.ф-лы, 5 ил., 2 табл. , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , ОПИСАНИЕ ИЗОБРЕТЕНИЯ К ПАТЕНТУИзобретение относится к фрагментам гибридного токсина и содержащим эти фрагменты токсинам, полученным из инсектицидных кристаллических белков Bacillus thuringiensis. Bacillus thuringiensis(далее B. t. ) способна продуцировать белки, накапливаемые внутри клеток в виде кристаллов. Эти кристаллические белки токсичны для личинок многих насекомых. Кристаллические белки подразделяют на различные классы на основе гомологических рядов и типов токсичности. Наиболее изучен CryI-класс белков, который представлен протоксинами с молекулярной массой 140 кДа и эффективен против лепидоптер. В некоторой степени принцип действия кристаллических белков объясним. После перорального приема кристаллы растворяются в щелочной среде средней кишки личинки. Растворенные белки затем под действием пищеварительной протеиназы превращаются в устойчивый к протеиназе токсичный фрагмент с молекулярной массой приблизительно 65 кДа, который связывается с рецепторами эпителиальных клеток в средней кишке насекомого и проникает сквозь мембраны клеток. Это приводит к разрыву клеток и гибели личинки. Спектр действия конкретного кристаллического белка во многом зависит от наличия рецепторов на эпителиальных клетках пищеварительного тракта пораженных насекомых. Указанный спектр совместно обусловлен растворимостью кристаллического белка и его протеолитической активацией in vivo. Важность связывания кристаллического белка с пищеварительными эпителиальными рецепторами дополнительно подтверждается тем, что у насекомых, характеризуемых устойчивостью к одному из кристаллических белков, связи кристаллических белков с эпителиальными клетками средней кишки существенно ослаблены. Считается, что токсичные фрагменты кристаллических белков состоят из трех определенных структурных доменов. Домен I, крайний N-терминальный, состоит из 7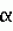 -спиралей. Домен II содержит 3 -спиралей. Домен II содержит 3  -складки, а домен III (крайний С-терминальный) складчато сложен (свернут) в -слоистую структуру. В проекции на Cryl-последовательности домен I занимает положения аминокислотных остатков с 28 по 260, домен II приблизительно с 260 по 460 и домен III приблизительно с 460 по 600. Предлагаемое изобретение относится, в частности, но не исключительно, к гибридным кристаллическим белкам, включая CryIC и CryIE или CryIA. Последовательность (далее этот термин обозначается SEQ с соответствующими латинскими индексами) нуклеотидов CrylC-гена B.t. подвид entomocidus 60.5 приведены в SEQ ID No.1, а соответствующая последовательность аминокислот белка, кодированная указанной последовательностью нуклеотидов, приведена в SEQ ID No. 2. Последовательность нуклеотидов CryIE-гена B.t. подвид kenyae 4FI приведена в SEQ ID 3, а соответствующая последовательность аминокислот белка, кодированная указанной последовательностью нуклеотидов, приведена в SEQ ID No.4. Эти белки токсичны для лепидоптер, но внутри этого вида насекомых каждый белок имеет разную специфичность. В частности, CryIC эффективен против S.exigua и M.brassicae. Согласно изобретению предложен фрагмент гибридного токсина B.t, содержащий на его С-конце домен III первого Сrу-белка, или часть этого домена, или белок, существенно сходный с этим доменом, при условии, что N-терминальная область фрагмента представлена N-терминальной областью второго Сrу-белка, или частью области, или белком, существенно сходным с этой областью. Предпочтительный фрагмент - это фрагмент, который не связывается с сайтом связывания CryIC во внутренностях насекомого, когда этот фрагмент содержит на его С-конце домен III белка CryIC, или часть этого домена, или белок, существенно сходный с этим доменом, или фрагмент, который не связывается с сайтом связывания белка CryIA, когда этот фрагмент содержит на своем С-конце домен III белка CryIA, или часть домена, или белок, существенно сходный с этим доменом. Термин "существенно сходный" относится к чистым белкам, имеющим аминокислотную последовательность, по меньшей мере на 75% сходную с последовательностью белков согласно изобретению. Предпочтительна степень сходства не менее на 85%, более предпочтительна - не менее 90% и еще более предпочтительна - не менее 95%. В контексте предлагаемого изобретения две аминокислотные последовательности, которые по меньшей мере на 75, 85, 90 и 95% сходны одна с другой, имеют по меньшей мере 75, 85, 90 и 95% идентичных или консервативно замененных аминокислотных остатков в тех же положениях при их оптимальном введении в цепь с образованием до 6 брешей при условии, что образующиеся бреши в общем не затрагивают более 15 остатков аминокислот. Консервативную замену аминокислот в соответствии с целью изобретения можно произвести в следующих группах: (I) серин и треонин, (II) глутаминовая кислота и аспарагиновая кислота, (III) аргинин и лизин, (IV) аспарагин и глутамин, (V) изолейцин, лейцин, валин и метионин, (VI) фенилаланин, тирозин и триптофан, (VII) аланин и глицин, при условии, что в SEQ ID No.6 консервативно заменены в положении 620 Ser и Туr, а Аlа и Glu - в положении 618, и что в SEQ ID No.8 Ser и Туr консервативно заменены в положении 627, а Аlа и Glu - в положении 625. Термин "часть" белка обозначает пептид, образованный указанным белком и содержащий не менее 80% его послеловательности. Термин "сайт связывания" обозначает участок молекулы, с которым токсин образует обратимую связь при Ка между ними порядка по меньшей мере 104 дм3 -складки, а домен III (крайний С-терминальный) складчато сложен (свернут) в -слоистую структуру. В проекции на Cryl-последовательности домен I занимает положения аминокислотных остатков с 28 по 260, домен II приблизительно с 260 по 460 и домен III приблизительно с 460 по 600. Предлагаемое изобретение относится, в частности, но не исключительно, к гибридным кристаллическим белкам, включая CryIC и CryIE или CryIA. Последовательность (далее этот термин обозначается SEQ с соответствующими латинскими индексами) нуклеотидов CrylC-гена B.t. подвид entomocidus 60.5 приведены в SEQ ID No.1, а соответствующая последовательность аминокислот белка, кодированная указанной последовательностью нуклеотидов, приведена в SEQ ID No. 2. Последовательность нуклеотидов CryIE-гена B.t. подвид kenyae 4FI приведена в SEQ ID 3, а соответствующая последовательность аминокислот белка, кодированная указанной последовательностью нуклеотидов, приведена в SEQ ID No.4. Эти белки токсичны для лепидоптер, но внутри этого вида насекомых каждый белок имеет разную специфичность. В частности, CryIC эффективен против S.exigua и M.brassicae. Согласно изобретению предложен фрагмент гибридного токсина B.t, содержащий на его С-конце домен III первого Сrу-белка, или часть этого домена, или белок, существенно сходный с этим доменом, при условии, что N-терминальная область фрагмента представлена N-терминальной областью второго Сrу-белка, или частью области, или белком, существенно сходным с этой областью. Предпочтительный фрагмент - это фрагмент, который не связывается с сайтом связывания CryIC во внутренностях насекомого, когда этот фрагмент содержит на его С-конце домен III белка CryIC, или часть этого домена, или белок, существенно сходный с этим доменом, или фрагмент, который не связывается с сайтом связывания белка CryIA, когда этот фрагмент содержит на своем С-конце домен III белка CryIA, или часть домена, или белок, существенно сходный с этим доменом. Термин "существенно сходный" относится к чистым белкам, имеющим аминокислотную последовательность, по меньшей мере на 75% сходную с последовательностью белков согласно изобретению. Предпочтительна степень сходства не менее на 85%, более предпочтительна - не менее 90% и еще более предпочтительна - не менее 95%. В контексте предлагаемого изобретения две аминокислотные последовательности, которые по меньшей мере на 75, 85, 90 и 95% сходны одна с другой, имеют по меньшей мере 75, 85, 90 и 95% идентичных или консервативно замененных аминокислотных остатков в тех же положениях при их оптимальном введении в цепь с образованием до 6 брешей при условии, что образующиеся бреши в общем не затрагивают более 15 остатков аминокислот. Консервативную замену аминокислот в соответствии с целью изобретения можно произвести в следующих группах: (I) серин и треонин, (II) глутаминовая кислота и аспарагиновая кислота, (III) аргинин и лизин, (IV) аспарагин и глутамин, (V) изолейцин, лейцин, валин и метионин, (VI) фенилаланин, тирозин и триптофан, (VII) аланин и глицин, при условии, что в SEQ ID No.6 консервативно заменены в положении 620 Ser и Туr, а Аlа и Glu - в положении 618, и что в SEQ ID No.8 Ser и Туr консервативно заменены в положении 627, а Аlа и Glu - в положении 625. Термин "часть" белка обозначает пептид, образованный указанным белком и содержащий не менее 80% его послеловательности. Термин "сайт связывания" обозначает участок молекулы, с которым токсин образует обратимую связь при Ка между ними порядка по меньшей мере 104 дм3 моль-1. Фрагмент токсина может содержать на своем N-конце N-терминальную область любых инсектицидных белков B.t., известных как "Cry" или "Cyt", среди которых CryIA(a), CryIA(b), CryIA(c), CryIB, CryIC, CryID, CryIE, CryIF, CryIG, CryIH, CryIIA, CryIIB, CryIIC, CryIIIA, CryIIIB, CryIIIB(b), CryIVA, CryIVB, CryIVC, CryIVD, CYTA, CryX1(IIIC), CryX2(IIID), СrуХ3, CryV и СrуХ4, или часть этой области, или белок, существенно сходный с этой областью. Он также может содержать на своем С-конце домен III белка CryIC или часть домена, или белок, существенно сходный с этим доменом. Таким образом, фрагмент может включать домен II белка CryIE, CryIB, CryID или CryIA, или часть домена, или белок, существенно сходный с этим доменом, и домен III белка CryIC или часть этого домена III, или белок, существенно сходный с доменом III. В частности, предпочтительно, чтобы фрагмент содержал домены I и II белков CryIE, CryIB, CryID или CryIA, или их часть, или белок, существенно сходный с этими доменами, и домен III белка CryIC или его часть, или белок, существенно сходный с доменом III. Наиболее предпочтительно, чтобы токсичный фрагмент имел на своем С-конце область, содержащую последовательность аминокислот от 454 до 602 белка CrylC, или последовательность, существенно сходную с указанной последовательностью. Фрагмент может иметь на своем C-конце область, содержащую последовательность аминокислот от 478 до 602 белка CryIC, или последовательность, существенно сходную с указанной последовательностью, при условии, что непосредственное объединение последовательности, включающей аминокислоты от 478 до 602 белка CryIC, с С-концом домена II белков CryIA, CryIB, CryIC, CryID, CryIE, даст продукт слияния, отвечающий условиям получения инсектицидного компонента фрагмента. Специалисту понятно, что может возникнуть необходимость добавить пептидную область к С-концу домена II, которая отдаляет С-терминальную область белка CrylC, позволяя ей уложиться таким образом, чтобы проявилась инсектицидная активность. Особенно предпочтительно, чтобы токсичный фрагмент согласно изобретению содержал I) последовательность аминокислот приблизительно от 1 до примерно 620 в SEQ ID No.6 или последовательность аминокислот приблизительно от 1 до приблизительно 620 в SEQ ID No.6, в которых произведена по меньшей мере одна из следующих замен: Ilе в положении 609 замещен Leu; Ala в положении 618 замещен Glu; Ser в положении 620 замещен Туr, или II) последовательность аминокислот от приблизительно 1 до примерно 627 в SEQ ID No. 8 или последовательность аминокислот от приблизительно 1 до приблизительно 627 в SEQ ID No.8, в которых произведена по меньшей мере одна из следующих замен: Ilе в положении 616 замещен Leu; Ala в положении 625 замещен Glu; Ser в положении 627 замещен Туr. Какими бы ни были допустимые замены аминокислот, один или более из следующих остатков (обозначения последовательности даны согласно последовательности белка CryIC) не подлежат изменению: Phe(501); Val(478); Trp(479) и Thr(486). Изобретение также относится к гибридному токсину, содержащему указанный фрагмент или токсин по меньшей мере на 85% сходный с таким гибридным токсином, который имеет существенно близкую инсектицидную активность или может связывать рецепторы. Кроме того, изобретение относится к чистым белкам, которые по меньшей мере на 90% идентичны фрагментам токсина или гибридным токсинам согласно изобретению. Изобретение относится и к рекомбинантной ДНК, содержащей последовательность, кодирующую белок, имеющий последовательность аминокислот описанных выше токсинов или их фрагментов. Изобретение также относится к рекомбинантной ДНК, содержащей последовательность от нуклеотида 1 до нуклеотида 1860 в SEQ ID No.5 или к ДНК, сходной с ней, кодирующей существенно сходный белок, или к рекомбинантной ДНК, содержащей последовательность от нуклеотида 1 до нуклеотида 1881 в SEQ ID Nо.7, или к ДНК, сходной с ней, кодирующей существенно сходный белок. Термин "сходная ДНК" означает, что интересующая последовательность способна образовывать гибрид с рекомбинантной последовательностью согласно изобретению. Когда интересующая последовательность и последовательность по изобретению являются двунитевыми, нуклеиновая кислота, образующая интересующую последовательность, имеет ТМ в пределах 20oС от ТМ последовательности по изобретению. В том случае, когда интересующую последовательность и последовательность по изобретению смешивают и одновременно денатурируют, значения ТМ этих последовательностей находятся предпочтительно в пределах 10oС одна по отношению к другой. Более предпочтительно гибридизацию проводят в жестких условиях, причем предпочтительно фиксируют интересующую ДНК или ДНК по изобретению. Таким образом, денатурированную либо интересующую, либо последовательность по изобретению предпочтительно сначала связывают с подложкой и проводят гибридизацию в течение определенного периода времени при температуре 50-70oС в двукратном цитратном буферном солевом растворе, содержащем 0,1% додецилсульфата натрия, после чего подложку промывают при той же температуре, но в буферном растворе с пониженной концентрацией цитрата натрия. В зависимости от необходимой жесткости условий и, следовательно, степени сходства последовательностей обычно выбирают следующие буферные растворы: однократный цитрат натрия, содержащий 0,1% додецилсульфата натрия, двукратно разбавленный цитрат натрия, содержащий 0,1% додецилсульфата натрия, десятикратно разбавленный цитрат натрия, содержащий 0,1% додецилсульфата натрия. К последовательностям, имеющим наивысшую степень сходства, относятся такие, на гибридизацию которых менее всего влияет промывка в буферных растворах пониженной концентрации. Наиболее предпочтительно, чтобы интересующая последовательность и последовательность по изобретению были сходны настолько, что промывка или инкубация в десятикратно разбавленном буферном растворе цитрата натрия, содержащем 0,1% додецилсульфата натрия, не могла существенно повлиять на их гибридизацию. Рекомбинантная ДНК может также кодировать белок, обладающий гербицидной устойчивостью, стимулирующий рост растений, обладающий фунгицидными, бактерицидными, антивирусными и/или антинематодными свойствами. В случае, когда ДНК требуется ввести в гетерологичный организм, ее модифицируют, чтобы исключить известные основные элементы нестабильности мРНК (например, АТ-богатые области) и сигналы полиаденилирования, и/или используют кодоны, предпочтительные для организма, которому эта рекомбинантная ДНК предназначена, так что экспрессией такой модифицированной ДНК в указанном организме синтезируется белок, по существу сходный с белком, синтезируемым при экспрессии немодифицированной рекомбинантной ДНК в организме, для которого белковые компоненты гибридного токсина или фрагментов токсина являются эндогенными. Кроме того, изобретение относится к последовательности ДНК, комплементарной такой последовательности, которая гибридизирована в жестких условиях с рекомбинантной ДНК согласно изобретению. Изобретение также касается вектора, содержащего такую рекомбинантную (или комплементарную ей) последовательность ДНК; растения или микроорганизма, которые включают и в которых возможна экспрессия ДНК; растений, трансформированных с помощью такой ДНК; потомства таких растений, которое содержит ДНК, устойчиво внедренную и способную нести наследственные признаки по Менделю, и/или семян таких растений и такого потомства. Кроме того, изобретение относится к белку, полученному экспрессией указанной ДНК, и инсектицидному белку, полученному экспрессией рекомбинантной ДНК в растениях, трансформированных этим белком. Кроме того, изобретение относится к инсектицидной композиции, содержащей один или более токсичных фрагментов или содержащих их токсинов согласно изобретению; к способу борьбы с насекомыми, который заключается в обработке их такими фрагментами или токсинами, или композициями, и способу экстракции для получения инсектицидных белков из содержащего их органического материала, заключающемуся в том, что этот материал вымачивают и экстрагируют растворителем. Далее изобретение будет описано на примере получения фрагментов гибридных токсинов B.t. согласно изобретению со ссылками на соответствующие чертежи и перечень последовательностей. На фиг. 1 показана схема образования генов гибридного кристаллического белка через рекомбинацию in vivo. Образованы тандемные плазмиды (pBD560 и pBD 650), несущие два укороченных гена кристаллического белка, имеющих ориентацию с прямым поворотом. 5"-ген (контурный прямоугольник), не имеет области, кодирующей протоксин (сплошной прямоугольник), а у 3"-гена (заштрихованный прямоугольник) часть области, кодирующей домен I, удалена. Рекомбинация in vivo между гомологичными областями (домены II и III) имеет место в recA + штамм JM101. Отбор рекомбинантов путем гидролитического расщепления (нерекомбинантов) с помощью NotI и BamHI с последующей трансформацией дает ряд плазмид, кодирующих гибридные кристаллические белки. На фиг. 2 показано расположение остатков аминокислот в положениях от 420 до 630 белков CryIE и CryIC. Граница между доменами II и III обозначена. Показаны только те аминокислотные остатки белков CryIC, которые отличаются от белка CryIE, одинаковые остатки обозначены пунктиром. Положения кроссовера (G27, Н13, Н7, Н8, Н17 и Н21) в фрагментах гибридного токсина белков CryIE-CryIC согласно изобретению также показаны. На фиг. 3 показано расположение остатков аминокислот в положениях от 420 до 630 белков CryIE и CryIC. Граница между доменами II и III обозначена. Показаны только те аминокислотные остатки белков GryIC, которые отличаются от белка CryIE, одинаковые остатки обозначены пунктиром. Положения кроссовера (F59, F71, F26 и Е7) в фрагментах гибридного токсина белков CryIE-CryIC согласно изобретению также показаны. На фиг. 4 показаны результаты некоторых экспериментов на гетерологическую конкуренцию. Биотинилированный белок CryIC (табло А) и G27 (табло В) инкубируют с везикулами S. exigua BBMV в отсутствие (цепочка а) или в присутствии избытка немеченого белка, как показано на табло. После инкубации везикулы промывают, загружают в полиакриламидный гель с додецилсульфатом натрия и блотируют на нитроцеллюлозные мембраны. Биотинилированные кристаллические белки, повторно отделенные с везикулами, визуализируют конъюгатом стрептавидин-пероксидаза. На фиг.4 они помечены стрелкой. На фиг. 5 показана карта плазмиды рSВ456, которая кодирует фрагмент гибридного токсина G27 и используется для трансформации штамма B.t.51, не вырабатывающего кристалический у токсин. SEQ ID No. 1 показывает последовательность нуклеотидсв CryIC-гена из B. t. подвид entomocidus 60.5. SEQ ID No. 2 показывает последовательность аминокислот белка, кодированного CryIC-геном, имеющимся в SEQ ID No.1. SEQ ID No. 3 показывает последовательность нуклеотидов CryIC-гена из B. t. подвид kenyae 4FI. SEQ ID No. 4 показывает последовательность аминокислот белка, кодированного CryIC-геном, показанным в SEQ ID No.3. SEQ ID No. 5 показывает последовательность нуклеотидов, кодирующую предпочтительный фрагмент гибридного токсина CryIE/CryIC B.t. согласно изобретению. SEQ ID No. 6 показывает последовательность аминокислот белка, кодированного последовательностью нуклеотидов, показанной в SEQ ID No.5. SEQ ID No. 7 показывает последовательность нуклеотидов фрагмента гибридного токсина CryIA/CryIC согласно изобретению. SEQ ID No. 8 показывает последовательность аминокислот белка, кодированного последовательностью нуклеотидов согласно SEQ ID No.7. Получение плазмид, кодирующих фрагменты гибридного токсина. При получении плазмид, несущих CryIC или CryIE гены, используют Escherichia coli XLI-blue (Stratagene Inc.) в качестве хозяина, исключая тот случай, когда штамм JM101 используют как rесА+фон. Вектор для экспрессии кристаллических белков в E.coli получают из рКК233-2 (Pharmacia LKB Biotechnology). Размер рКК233-2 уменьшают, удаляя EcoRI-PvuII фрагмент, несущий ген, кодирующий устойчивость к тетрациклину. Впоследствии линкер XhoI с шестью парами оснований встраивают в сайт HindIII, получая pBD10. Плазмиду ВК+ образуют, встраивая линкер BglII в сайт Sacl плазмиды Bluescript SK+ (Stratagene Inc. ). Полилинкер плазмиды ВК+ на участке от BglII до XhoI встраивают в сайт NcoI-XhoI в pBD10. Полученный экспрессирующий вектор pBD11 содержит ярко выраженный trc промотор, lacZ сайт связывания рибосом и инициирующий ATG кодон. Инициирующий кодон частично перекрывается сайтом Ncol и за ним следует полилинкер, что облегчает включение вставок в вектор. Транскрипция заканчивается rrnВ терминатором транскрипции. Гены сrуIС и cryIE клонируют соответственно из B.t. подвид entomocidus 60.5 и kenya 4F1, как ранее описано Honee et al., 1990 (Appl. Environ. Microbiol. , 56, р. 823-825); Visser et al., 1990 (J. Bacteriol., 172, р. 6783-6788). Для клонирования мутагенезом in vitro был создан сайт NсоI, частично перекрывающийся с инициирующим кодоном гена cryIC. Сайт BglII был создан непосредственно за терминирующим кодоном белка cryIC сайт-специфическим мутагенезом, в результате чего была получена последовательность моль-1. Фрагмент токсина может содержать на своем N-конце N-терминальную область любых инсектицидных белков B.t., известных как "Cry" или "Cyt", среди которых CryIA(a), CryIA(b), CryIA(c), CryIB, CryIC, CryID, CryIE, CryIF, CryIG, CryIH, CryIIA, CryIIB, CryIIC, CryIIIA, CryIIIB, CryIIIB(b), CryIVA, CryIVB, CryIVC, CryIVD, CYTA, CryX1(IIIC), CryX2(IIID), СrуХ3, CryV и СrуХ4, или часть этой области, или белок, существенно сходный с этой областью. Он также может содержать на своем С-конце домен III белка CryIC или часть домена, или белок, существенно сходный с этим доменом. Таким образом, фрагмент может включать домен II белка CryIE, CryIB, CryID или CryIA, или часть домена, или белок, существенно сходный с этим доменом, и домен III белка CryIC или часть этого домена III, или белок, существенно сходный с доменом III. В частности, предпочтительно, чтобы фрагмент содержал домены I и II белков CryIE, CryIB, CryID или CryIA, или их часть, или белок, существенно сходный с этими доменами, и домен III белка CryIC или его часть, или белок, существенно сходный с доменом III. Наиболее предпочтительно, чтобы токсичный фрагмент имел на своем С-конце область, содержащую последовательность аминокислот от 454 до 602 белка CrylC, или последовательность, существенно сходную с указанной последовательностью. Фрагмент может иметь на своем C-конце область, содержащую последовательность аминокислот от 478 до 602 белка CryIC, или последовательность, существенно сходную с указанной последовательностью, при условии, что непосредственное объединение последовательности, включающей аминокислоты от 478 до 602 белка CryIC, с С-концом домена II белков CryIA, CryIB, CryIC, CryID, CryIE, даст продукт слияния, отвечающий условиям получения инсектицидного компонента фрагмента. Специалисту понятно, что может возникнуть необходимость добавить пептидную область к С-концу домена II, которая отдаляет С-терминальную область белка CrylC, позволяя ей уложиться таким образом, чтобы проявилась инсектицидная активность. Особенно предпочтительно, чтобы токсичный фрагмент согласно изобретению содержал I) последовательность аминокислот приблизительно от 1 до примерно 620 в SEQ ID No.6 или последовательность аминокислот приблизительно от 1 до приблизительно 620 в SEQ ID No.6, в которых произведена по меньшей мере одна из следующих замен: Ilе в положении 609 замещен Leu; Ala в положении 618 замещен Glu; Ser в положении 620 замещен Туr, или II) последовательность аминокислот от приблизительно 1 до примерно 627 в SEQ ID No. 8 или последовательность аминокислот от приблизительно 1 до приблизительно 627 в SEQ ID No.8, в которых произведена по меньшей мере одна из следующих замен: Ilе в положении 616 замещен Leu; Ala в положении 625 замещен Glu; Ser в положении 627 замещен Туr. Какими бы ни были допустимые замены аминокислот, один или более из следующих остатков (обозначения последовательности даны согласно последовательности белка CryIC) не подлежат изменению: Phe(501); Val(478); Trp(479) и Thr(486). Изобретение также относится к гибридному токсину, содержащему указанный фрагмент или токсин по меньшей мере на 85% сходный с таким гибридным токсином, который имеет существенно близкую инсектицидную активность или может связывать рецепторы. Кроме того, изобретение относится к чистым белкам, которые по меньшей мере на 90% идентичны фрагментам токсина или гибридным токсинам согласно изобретению. Изобретение относится и к рекомбинантной ДНК, содержащей последовательность, кодирующую белок, имеющий последовательность аминокислот описанных выше токсинов или их фрагментов. Изобретение также относится к рекомбинантной ДНК, содержащей последовательность от нуклеотида 1 до нуклеотида 1860 в SEQ ID No.5 или к ДНК, сходной с ней, кодирующей существенно сходный белок, или к рекомбинантной ДНК, содержащей последовательность от нуклеотида 1 до нуклеотида 1881 в SEQ ID Nо.7, или к ДНК, сходной с ней, кодирующей существенно сходный белок. Термин "сходная ДНК" означает, что интересующая последовательность способна образовывать гибрид с рекомбинантной последовательностью согласно изобретению. Когда интересующая последовательность и последовательность по изобретению являются двунитевыми, нуклеиновая кислота, образующая интересующую последовательность, имеет ТМ в пределах 20oС от ТМ последовательности по изобретению. В том случае, когда интересующую последовательность и последовательность по изобретению смешивают и одновременно денатурируют, значения ТМ этих последовательностей находятся предпочтительно в пределах 10oС одна по отношению к другой. Более предпочтительно гибридизацию проводят в жестких условиях, причем предпочтительно фиксируют интересующую ДНК или ДНК по изобретению. Таким образом, денатурированную либо интересующую, либо последовательность по изобретению предпочтительно сначала связывают с подложкой и проводят гибридизацию в течение определенного периода времени при температуре 50-70oС в двукратном цитратном буферном солевом растворе, содержащем 0,1% додецилсульфата натрия, после чего подложку промывают при той же температуре, но в буферном растворе с пониженной концентрацией цитрата натрия. В зависимости от необходимой жесткости условий и, следовательно, степени сходства последовательностей обычно выбирают следующие буферные растворы: однократный цитрат натрия, содержащий 0,1% додецилсульфата натрия, двукратно разбавленный цитрат натрия, содержащий 0,1% додецилсульфата натрия, десятикратно разбавленный цитрат натрия, содержащий 0,1% додецилсульфата натрия. К последовательностям, имеющим наивысшую степень сходства, относятся такие, на гибридизацию которых менее всего влияет промывка в буферных растворах пониженной концентрации. Наиболее предпочтительно, чтобы интересующая последовательность и последовательность по изобретению были сходны настолько, что промывка или инкубация в десятикратно разбавленном буферном растворе цитрата натрия, содержащем 0,1% додецилсульфата натрия, не могла существенно повлиять на их гибридизацию. Рекомбинантная ДНК может также кодировать белок, обладающий гербицидной устойчивостью, стимулирующий рост растений, обладающий фунгицидными, бактерицидными, антивирусными и/или антинематодными свойствами. В случае, когда ДНК требуется ввести в гетерологичный организм, ее модифицируют, чтобы исключить известные основные элементы нестабильности мРНК (например, АТ-богатые области) и сигналы полиаденилирования, и/или используют кодоны, предпочтительные для организма, которому эта рекомбинантная ДНК предназначена, так что экспрессией такой модифицированной ДНК в указанном организме синтезируется белок, по существу сходный с белком, синтезируемым при экспрессии немодифицированной рекомбинантной ДНК в организме, для которого белковые компоненты гибридного токсина или фрагментов токсина являются эндогенными. Кроме того, изобретение относится к последовательности ДНК, комплементарной такой последовательности, которая гибридизирована в жестких условиях с рекомбинантной ДНК согласно изобретению. Изобретение также касается вектора, содержащего такую рекомбинантную (или комплементарную ей) последовательность ДНК; растения или микроорганизма, которые включают и в которых возможна экспрессия ДНК; растений, трансформированных с помощью такой ДНК; потомства таких растений, которое содержит ДНК, устойчиво внедренную и способную нести наследственные признаки по Менделю, и/или семян таких растений и такого потомства. Кроме того, изобретение относится к белку, полученному экспрессией указанной ДНК, и инсектицидному белку, полученному экспрессией рекомбинантной ДНК в растениях, трансформированных этим белком. Кроме того, изобретение относится к инсектицидной композиции, содержащей один или более токсичных фрагментов или содержащих их токсинов согласно изобретению; к способу борьбы с насекомыми, который заключается в обработке их такими фрагментами или токсинами, или композициями, и способу экстракции для получения инсектицидных белков из содержащего их органического материала, заключающемуся в том, что этот материал вымачивают и экстрагируют растворителем. Далее изобретение будет описано на примере получения фрагментов гибридных токсинов B.t. согласно изобретению со ссылками на соответствующие чертежи и перечень последовательностей. На фиг. 1 показана схема образования генов гибридного кристаллического белка через рекомбинацию in vivo. Образованы тандемные плазмиды (pBD560 и pBD 650), несущие два укороченных гена кристаллического белка, имеющих ориентацию с прямым поворотом. 5"-ген (контурный прямоугольник), не имеет области, кодирующей протоксин (сплошной прямоугольник), а у 3"-гена (заштрихованный прямоугольник) часть области, кодирующей домен I, удалена. Рекомбинация in vivo между гомологичными областями (домены II и III) имеет место в recA + штамм JM101. Отбор рекомбинантов путем гидролитического расщепления (нерекомбинантов) с помощью NotI и BamHI с последующей трансформацией дает ряд плазмид, кодирующих гибридные кристаллические белки. На фиг. 2 показано расположение остатков аминокислот в положениях от 420 до 630 белков CryIE и CryIC. Граница между доменами II и III обозначена. Показаны только те аминокислотные остатки белков CryIC, которые отличаются от белка CryIE, одинаковые остатки обозначены пунктиром. Положения кроссовера (G27, Н13, Н7, Н8, Н17 и Н21) в фрагментах гибридного токсина белков CryIE-CryIC согласно изобретению также показаны. На фиг. 3 показано расположение остатков аминокислот в положениях от 420 до 630 белков CryIE и CryIC. Граница между доменами II и III обозначена. Показаны только те аминокислотные остатки белков GryIC, которые отличаются от белка CryIE, одинаковые остатки обозначены пунктиром. Положения кроссовера (F59, F71, F26 и Е7) в фрагментах гибридного токсина белков CryIE-CryIC согласно изобретению также показаны. На фиг. 4 показаны результаты некоторых экспериментов на гетерологическую конкуренцию. Биотинилированный белок CryIC (табло А) и G27 (табло В) инкубируют с везикулами S. exigua BBMV в отсутствие (цепочка а) или в присутствии избытка немеченого белка, как показано на табло. После инкубации везикулы промывают, загружают в полиакриламидный гель с додецилсульфатом натрия и блотируют на нитроцеллюлозные мембраны. Биотинилированные кристаллические белки, повторно отделенные с везикулами, визуализируют конъюгатом стрептавидин-пероксидаза. На фиг.4 они помечены стрелкой. На фиг. 5 показана карта плазмиды рSВ456, которая кодирует фрагмент гибридного токсина G27 и используется для трансформации штамма B.t.51, не вырабатывающего кристалический у токсин. SEQ ID No. 1 показывает последовательность нуклеотидсв CryIC-гена из B. t. подвид entomocidus 60.5. SEQ ID No. 2 показывает последовательность аминокислот белка, кодированного CryIC-геном, имеющимся в SEQ ID No.1. SEQ ID No. 3 показывает последовательность нуклеотидов CryIC-гена из B. t. подвид kenyae 4FI. SEQ ID No. 4 показывает последовательность аминокислот белка, кодированного CryIC-геном, показанным в SEQ ID No.3. SEQ ID No. 5 показывает последовательность нуклеотидов, кодирующую предпочтительный фрагмент гибридного токсина CryIE/CryIC B.t. согласно изобретению. SEQ ID No. 6 показывает последовательность аминокислот белка, кодированного последовательностью нуклеотидов, показанной в SEQ ID No.5. SEQ ID No. 7 показывает последовательность нуклеотидов фрагмента гибридного токсина CryIA/CryIC согласно изобретению. SEQ ID No. 8 показывает последовательность аминокислот белка, кодированного последовательностью нуклеотидов согласно SEQ ID No.7. Получение плазмид, кодирующих фрагменты гибридного токсина. При получении плазмид, несущих CryIC или CryIE гены, используют Escherichia coli XLI-blue (Stratagene Inc.) в качестве хозяина, исключая тот случай, когда штамм JM101 используют как rесА+фон. Вектор для экспрессии кристаллических белков в E.coli получают из рКК233-2 (Pharmacia LKB Biotechnology). Размер рКК233-2 уменьшают, удаляя EcoRI-PvuII фрагмент, несущий ген, кодирующий устойчивость к тетрациклину. Впоследствии линкер XhoI с шестью парами оснований встраивают в сайт HindIII, получая pBD10. Плазмиду ВК+ образуют, встраивая линкер BglII в сайт Sacl плазмиды Bluescript SK+ (Stratagene Inc. ). Полилинкер плазмиды ВК+ на участке от BglII до XhoI встраивают в сайт NcoI-XhoI в pBD10. Полученный экспрессирующий вектор pBD11 содержит ярко выраженный trc промотор, lacZ сайт связывания рибосом и инициирующий ATG кодон. Инициирующий кодон частично перекрывается сайтом Ncol и за ним следует полилинкер, что облегчает включение вставок в вектор. Транскрипция заканчивается rrnВ терминатором транскрипции. Гены сrуIС и cryIE клонируют соответственно из B.t. подвид entomocidus 60.5 и kenya 4F1, как ранее описано Honee et al., 1990 (Appl. Environ. Microbiol. , 56, р. 823-825); Visser et al., 1990 (J. Bacteriol., 172, р. 6783-6788). Для клонирования мутагенезом in vitro был создан сайт NсоI, частично перекрывающийся с инициирующим кодоном гена cryIC. Сайт BglII был создан непосредственно за терминирующим кодоном белка cryIC сайт-специфическим мутагенезом, в результате чего была получена последовательность  (стоп-кодон подчеркнут). Фрагмент NcoI-BglII, содержащий cryIC кодирующую область, встраивают в вектор pBD11, получая плазмиду pBD150, дающую возможность экспрессии белка CryIC. Плазмида pBD155 является производной от pBD150, в которой последовательность 3" полилинкеров гена cryIC удалена. Фрагмент DraI плазмиды рЕМ14 (Visser et al., 1990), содержащий полный ген cryIE, клонируется в сайте EcoRV плазмиды SK+, в результате чего получается плазмида рЕМ15. Впоследствии сайт NcoI вводят сайт-специфическим мутагенезом в инициирующий кодон этого гена, и cryIE переносят как фрагмент NcoI-XhoI в вектор pBD11, получая плазмиду рВD60, дающую возможность экспрессии белка CryIE. Получены плазмиды, несущие только области, кодирующие токсичный фрагмент сrуI генов. Линкеры ВglII встроены в сайты ХmnI, представленные в паре оснований для положения 1835 гена cryIC, и в сайт HgiAI в положении 1839 гена cryIE. Впоследствии фрагменты Ncol-BglII, содержащие области, кодирующие токсичные фрагменты генов сrуIС (пара оснований 1835) и cryIE (пара оснований 1839), встраивают в вектор pBD11, получая соответственно плазмиды pBD151 и pBD161, как описано ниже. Тандемные плазмиды, используемые для получения гибридных генов cryIC-cryIE, конструируют следующим образом. Линкеры BamHI лигируют в плазмиде pBD160, гидролизованной с помощью НраI. Эту ДНК инкубируют с ВаmНI и XhoI, а укороченный ген cryIE, начинающийся от пары оснований 704, встраивают в плазмиду pBD151, получая плазмиду pBD560. Чтобы сконструировать тандемную плазмиду для получения гибридов сrуIЕ-сrуIС, плазмиду pBD155 гидролизуют с помощью NsiI и XhoI. Фрагмент, несущий укороченный ген cryIC, начинающийся от пары оснований 266, встраивают в плазмиду pBD161, гидролизованную PstI/XhoI, получая плазмиду pBD650. Присутствие уникальных сайтов рестрикции на NotI и BamHI между укороченными генами cryI в тандемных плазмидах pBD560 и pBD650 обусловлено последовательностями полилинкеров. Манипулирование ДНК и конструирование гибридного токсина. Все приемы манипулирования рекомбинантной ДНК проводили в соответствии с Sambrook et al.,1989 (в "Molecular Cloning, A Laboratory Manual: Cold Spring Harbour Press, Cold Spring Harbour), причем последовательность ДНК расшифровывали дидезокситрифосфатным методом с флуоресцентным окрашиванием дидезоксинуклеотидов, используя анализатор последовательности нуклеотидов Applied Biosystems 370A. Выявленная степень подобия cryI генов позволяет проводить внутримолекулярную рекомбинацию in vivo. В результате получают пару тандемных плазмид, каждая из которых несет два укороченных гена кристаллических белков, перекрывающиеся только в доменах II и III. Таким образом, рекомбинация происходит только в областях, кодирующих домены II и III. Рамочные рекомбинации, которые можно отбирать, используя рестриктазы для гидролиза, производят плазмиды, дающие возможность экспрессии полномерных гибридных протоксинов с молекулярной массой 140 кДа. Для получения in vivo рекомбинантов тандемную плазмиду (pBD560 или pBD650; фиг.2) переносят в JM101. ДНК в количестве 5 мкг выделяют из отдельно полученных рекомбинантов и гидролизуют с помощью NotI и BamHI, делая разрез между двумя укороченными cryI генами для селекции из нерекомбинантов, и ДНК трансформируется в E.coli XL1-голубую. Затем выращивают 5 отдельных колоний и анализируют характер белков и содержание плазмид. Гибридные токсины CryIC-CryIE и CryIE-CryIC получают с использованием соответственно тандемных плазмид pBD560 и pBD650, допуская их рекомбинацию в rесА+фон, выделяют ДНК, гидролизуют и переносят в штамм rесА, как описано выше. 100 колоний из 20 отдельных экспериментов анализируют электрофорезом в полиакриламидном геле с додецилсульфатом натрия. Из этих колоний 85% дают белок молекулярной массой 140 кДа, задаваемый рамочными рекомбинациями соответственно генов сrуIС и cryIE, cryIE и cryIC. E.coli продуцируют белки CryI в виде кристаллов, растворяющихся in vitro при высоких значениях рН. Приблизительно 15% гибридных токсинов, полученных описанным способом, растворяются при высоких значениях рН. Рекомбинанты, продуцирующие растворимые гибридные токсины, сначала классифицируют с использованием рестриктазы, затем определяют положение кроссовера в селекционированных гибридах каждого класса, анализируя последовательность ДНК. Все кроссоверы, которые приводят к получению растворимых гибридных токсинов, обнаруживаются очень близко от домена III или в нем. Очистка и анализ белков. Кристаллические белки выделяют в основном по методу Convents et al. (J. Biol. Chem. , 265, р.1369-1375; Eur. J. Biochem., 195, р.631-635). Вкратце, рекомбинантные E.coli выращивают при 30oС в 250 мл в ТВ-среде до ОП660 10-15. Кристаллы, выделенные из лизата E.coli, растворяют при инкубации в течение 2 часов в присутствии 20 мМоль Nа2СО3, 10 мМоль дитиотреитола, 100 мМоль NaCl при рН 10 и температуре 37oС. рН раствора понижают до 8 с помощью 100 мМоль буфера Tris-HCl, и инкубируют в присутствии трипсина. Раствор токсина диализируют против 20 мМоль буфера Tris-HCl и 100 мМоль NaCl при рН 9. Затем токсичный фрагмент очищают на колонке Mono Q 5/5, включенной в систему (FPLC) скоростной жидкостной хроматографии белков (Pharmacia LKB Biotechnology). Белки выделяют электрофорезом в полиакриламидном геле, содержащем 7,5% додецилсульфата натрия. Биохимический анализ и выделение токсичных фрагментов с молекулярной массой 65 кДа. Выделенные кристаллы очищенных белков CryIC, CryIE и гибридные белки растворяют при высоком значении рН и инкубируют с трипсином. Подобно белкам CryIC и СryIЕ все растворимые гибридные токсины при кроссовере в домене III превращаются в устойчивые фрагменты с молекулярной массой 65 кДа. Фрагменты с молекулярной массой 65 кДа можно очищать анионообменной хроматографией при таких же условиях, как и родительские белки. Tрипсин полностью разрушает гибриды F59 и F71, которые имеют кроссоверы в домене II. Несмотря на то, что эти гибриды не осаждаются в виде нерастворимых агрегаций, по-видимому, индуцированные трипсином места расщепления в родительских белках могут подвергнуться действию трипсина. Поэтому фрагменты с молекулярной массой 65 кДa не могут быть выделены из гибридов F59 и F71. В табл. 1 показана структура 5 гибридных токсинов CryIE-CryIC (G27, Н8, Н17, Н13, Н7) и 4 CryIC-CryIE гибридных токсинов (F59, F71, F26, Е7) в сравнении с белками CryIE и CryIC, из которых они получены. Аминокислотные последовательности гибридных токсинов CryIE-CryIC, содержащих токсичные фрагменты согласно изобретению, занимают положения до аминокислоты 1189 родительского белка CryIC, а гибридных токсинов CryIC-CryIE - до аминокислоты 1171 родительского белка CryIE. В табл. 1 приведены также значения относительной инсектицидной активности этих гибридных токсинов в сравнении с белками CryIE и CryIC. Большинство биологических анализов было проведено на очищенных фрагментах токсинов. Белки cryIC занимают положения от приблизительно аминокислоты 28 до 627, a cryIE - до 612. Полная длина последовательностей протоксинов указана в скобках. Определение токсичности для насекомых и инсектицидная активность cryICI cryIE продуктов гибридных генов. Бактериальные культуры концентрируют до ОП660 6,0 и 100 мкл наносят в виде точек на искусственную питательную среду на матрас для культуры тканей, имеющий 24 лунки. Наряду с этим разбавленные образцы очищенных токсинов наносят на питательную среду. Личинки во втором инстаре насекомых S.exigua, M. bras-sicae или M. sexta содержат на этой питательной среде (16-кратное разбавление на образец) 5 дней, после чего учитывают массу личинок. Относительный рост (равновесную концентрацию, снижающую рост на 50%) определяют по отношению средней массы личинок, содержащихся на питательной среде с токсином, к средней массе контрольных личинок, содержащихся на питательной среде без токсина. Насекомых вида M.sexta для кладки яиц поставляет Carolina Biological Supply Company, North Carolina, US. Токсичные фрагменты, кодированные продуктами гибридных генов, испытывают на активность против трех различных видов насекомых, описанных выше. Насекомые вида M. sexta восприимчивы как к CryIC, так и СrуIЕ. По их чувтствительности к трипсину можно предположить, что гибриды F59 и F71 не обладают активностью против этого насекомого (табл. 1). Несмотря на то, что Н7 конвертируется трипсином в устойчивые белки с молекулярной массой 65 кДа, он не токсичен для M. sexta. Все остальные гибриды, представленные в табл. 1, токсичны и, по-видимому, имеют нативную биологически активную конформацию. Фрагмент белка CrylC молекулярной массой 65 кДа в отличие от белка CryIE высокотоксичен по отношению к видам S. exigua и M.brassicae. Токсин G27 (табл. 1; фиг. 2), гибрид CryIE-CryIC с кроссовером в месте соединения доменов II и III активен по отношению к обоим видам насекомых. Это указывает на то, что домен III белка CryIC придает ему полную активность по отношению к S.exigua и M.brassicae. Гибрид Н8, который отличается от G27 только тремя аминокислотными остатками (см. фиг.3), будучи активным против M.sexta, не активен против S.exigua и M.brassicae. Антипод гибрида G27 - гибрид F26 (табл. 1, фиг.3), в котором домен III белка CryIC заменен доменом III белка CryIE, не активен против S.exigua или M. brassicae. Несмотря на то, что он токсичен по отношению к M.sexta, последовательности белка CryIC, включающие положения от аминокислот 28 до 462, вероятно, не достаточны для поражения S.exigua и M.brassicae. Последовательности белка CryIC только тогда восстановливают инсектицидную активность против этих насекомых, когда в этом гибриде (Е7) присутствуют аминокислотные остатки не ниже положения 602. В этом описании показано, что аминокислотные остатки в положениях 478-602 последовательности белка CrylC могут придать белку CrylE высокую инсектицидную активность против S.exigua и M.brassicae. Биотинилирование кристаллических белков и их анализ по степени связывания. Биотинилирование проводят с использованием биотин-N-гидроксисукцинимиидного эфира по существу таким образом, как оно описано изготовителем (Amersham). 1 мг кристаллического белка инкубируют в течение одного часа с 40 мкл биотинилирующего реагента в присутствии 50 мМоль NаНСО3, 15 мМоль NaCl при рН 8 и температуре 20oС. Раствор заливают в колонку, заполненную Sephadex 25, уравновешенным тем же буфером, содержащим 0,1% бычьего сывороточного альбумина для удаления несвязанного биотина, и образцы фракций наносят пятнами на нитроцеллюлозную мембрану. Фракции, содержащие биотинилированные кристаллические белки, визуализируют с помощью конъюгата стрептавидин-пероксидаза (Amersham), который катализирует окисление люминола, вызывающего хемилюминесценцию (ECL, Amersham), и сливают. Мембранные везикулы собирают по Wolfersberger et al. (Сотр. Biochem. Physiol., 86a, p.301-308, 1987) и, кроме того, еще раз промывают изолирующим буфером, содержащим 0,1% среды Tween-20. Связывание биотинилированных кристаллических белков с мембранными везикулами (100 мкг/мл) проводят в 100 мкл среды PBS, содержащей 1% бычьего сывороточного альбумина и 0,1% среды Tween-20 при рН7,6. Везикулы (20 мкг белка) инкубируют с 10 мг биотинилированных кристаллических белков в течение одного часа при 20oС в присутствии или при отсутствии 1000-кратного избытка немеченых кристаллических белков. Затем везикулы снова отделяют центрифугированием в течение 10 мин при 14000 g в центрифуге Eppendorf, дважды промывают связующим буфером, снова суспендируют в образцовом буфере, денатурируют нагреванием и вводят в 7,5%-ный полиакриламидный гель. После электрофореза белки блотируют на нитроцеллюлозные мембраны и биотинилированные кристаллические белки, которые повторно выделены с везикулами, визуализуют, инкубируя нитроцеллюлозу с конъюгатом стрептавидин-пероксидаза (Amersham), который катализирует окисление люминола, вызывающего хемилюминесценцию (ECL, Amersham). Поскольку связывание с эпителиальными клетками средней кишки - основная часть механизма действия кристаллических белков, их связывание мембранными везикулами S.exigna исследуют в экспериментах на гетерологическую конкуренцию. Конкурентные эксперименты показывают, что связывание меченого белка CryIC (фиг. 4А, цепочка а) и меченого гибрида F26 (не показан) может быть исключено при избытке немеченого белка CryIC (цепочка b) или гибрида F26 (цепочка е), но не при избытке гибрида G27 (цепочка с) или белка CryIE (цепочка d). Кроме того, связывание меченого гибрида G27 (фиг.4В, цепочка а) и меченого белка CryIE (не показан) может быть исключено при избытке гибрида G27 (цепочка b) или белка CryIE (цепочка d), но не при избытке белка CryIC (цепочка а) или гибрида F26 (цепочка е). Это позволяет сделать вывод, что G27 и CryIE "узнают" те же сайты связывания на мембранах средней кишки S. exigna и что эти сайты отличаются от сайтов, "узнаваемых" белком СrуIС и гибридом F26. Совместные испытания на токсичность и степень связывания показывают, что G27 обладает такой же токсичностью, как и CryIC, но связывается с иным рецептором. Поскольку насекомые могут приобретать сопротивляемость к кристаллическим белкам, изменяя показатели связывания рецепторов, G27 может быть полезен для контроля сопротивляемости насекомых в качестве варианта замены CryIC. Экспрессия генов гибридного токсина cryIE/cryIC в гетерологичных системах. Ген cryIE/cryIC гибридного токсина G27 экспрессируется в E.coli, и генный продукт проявляет по меньшей мере такую же инсектицидную активность (по меньшей мере против Spodoptera) как CryIC. Более того, этот продукт проявляет повышенную активность, если ген экспрессируется в штамме Bacillys thuringienses (см. ниже). Ген, кодирующий гибридный токсин G27, вводят в подходящий вектор-челнок, который затем вводят в соответствующего хозяина B.t. Такие трансформированные клетки культивируют, а получаемый от обеих цельных культур токсин и очищенные кристаллы проверяют на инсектицидную активность. Конструирование вектора-челнока, содержащего G27, трансформация штамма Bt 51 и очистка от него токсичного белка. Ген, кодирующий гибрид G27 (3400 пар оснований), отщепляют от плазмиды, индуцирующей экспрессию рКК233 Е.соli, с использованием Nсо1 и Хhо1. Сайт Xho1 заполняют фрагментом Кленова, т.е. фрагментом ДНК-полимеразы I от Е. соli. Полученный фрагмент лигируют с Nco1/Sma1, гидролизованной рSB635 (pBluescript KS+, PcryIC и CryIA(c) терминаторы транскрипции). Полученную плазмиду рSВ454 гидролизуют с Ара1 и Nоt1, образуя фрагмент с 4200 пар оснований, несущий промотор, гибрид G27 и терминатор. Этот фрагмент лигируют с Ара1/Not1 гидролизованной рSВ634 (челнок-вектор, содержащий рВС16.1 и pBluescript KS+), образуя рSВ456 (см. фиг.5). Плазмиду ДНК, выделенную из E.coli, ДН10В, используют для трансформации B.t.51, штамма B.t., не вырабатывающего кристаллический токсин. Положительные изоляты устойчивы к тетрациклину, что говорит о присутствии pSB456, и содержат большие включения, соответствующие белку с молекулярной массой 135 кДа (выявлено с помощью электрофореза в полиакриламидном геле с додецилсульфатом натрия). Образцы гибридного токсина G27 готовят из культур трансформированного штамма Bt51, выращенного спорообразованием при 30oС в среде цистеин-тетрациклин10. Биологический анализ инсектицидности (табл. 2) проводят как на полных цельных культурах, так и на отмытых препаратах кристаллического белка. Контрольные образцы включают Bt51 (рSВ440), содержащий токсин CryIC, и Bt51 (рSВ636), содержащий CryIE. Концентрацию токсина определяют электрофорезом в полиакриламидном геле с додецилсульфатом натрия. Несмотря на то, что изобретение описано на примере получения гибридного токсина G27, специалисту будет понятно, что, пользуясь информацией из этого описания, можно получить многие другие гибридные токсины с улучшенными инсектицидными показателями. Например, в SEQ ID No.7 и 8 представлены другие гибридные токсины согласно изобретению, которые обладают повышенной инсектицидной активностью. Можно получить гибридные токсины, содержащие домен III белка CryIC и N-конец любого иного белка Cry. Что касается биологических испытаний, триансформанты, несущие гибридный токсин, можно вырастить в SOP средах для ускорения испытаний и уменьшения объемов используемого материала. Кроме того, ген, кодирующий G27 и/или другие гибридные токсины, согласно изобретению можно перенести в кодирующие токсин штаммы B.t. и/или интегрировать в хромосому выведенных штамов B.t. или даже ввести в геномы растений для обеспечения инсектицидной активности in situ в этом растении. В этом случае особенно предпочтительно рекомбинантную ДНК, кодирующую токсины, модифицировать так, чтобы кодоны, предпочтительные для растения, которому эта рекомбинантная ДНК предназначена, были использованы так, что экспрессией такой модифицированной ДНК в указанном растении синтезируется белок, по существу сходный с белком, синтезируемым при экспрессии немодифицированной рекомбинантной ДНК в организме, для которого белковые компоненты гибридного токсина или фрагментов токсина являются эндогенными. (стоп-кодон подчеркнут). Фрагмент NcoI-BglII, содержащий cryIC кодирующую область, встраивают в вектор pBD11, получая плазмиду pBD150, дающую возможность экспрессии белка CryIC. Плазмида pBD155 является производной от pBD150, в которой последовательность 3" полилинкеров гена cryIC удалена. Фрагмент DraI плазмиды рЕМ14 (Visser et al., 1990), содержащий полный ген cryIE, клонируется в сайте EcoRV плазмиды SK+, в результате чего получается плазмида рЕМ15. Впоследствии сайт NcoI вводят сайт-специфическим мутагенезом в инициирующий кодон этого гена, и cryIE переносят как фрагмент NcoI-XhoI в вектор pBD11, получая плазмиду рВD60, дающую возможность экспрессии белка CryIE. Получены плазмиды, несущие только области, кодирующие токсичный фрагмент сrуI генов. Линкеры ВglII встроены в сайты ХmnI, представленные в паре оснований для положения 1835 гена cryIC, и в сайт HgiAI в положении 1839 гена cryIE. Впоследствии фрагменты Ncol-BglII, содержащие области, кодирующие токсичные фрагменты генов сrуIС (пара оснований 1835) и cryIE (пара оснований 1839), встраивают в вектор pBD11, получая соответственно плазмиды pBD151 и pBD161, как описано ниже. Тандемные плазмиды, используемые для получения гибридных генов cryIC-cryIE, конструируют следующим образом. Линкеры BamHI лигируют в плазмиде pBD160, гидролизованной с помощью НраI. Эту ДНК инкубируют с ВаmНI и XhoI, а укороченный ген cryIE, начинающийся от пары оснований 704, встраивают в плазмиду pBD151, получая плазмиду pBD560. Чтобы сконструировать тандемную плазмиду для получения гибридов сrуIЕ-сrуIС, плазмиду pBD155 гидролизуют с помощью NsiI и XhoI. Фрагмент, несущий укороченный ген cryIC, начинающийся от пары оснований 266, встраивают в плазмиду pBD161, гидролизованную PstI/XhoI, получая плазмиду pBD650. Присутствие уникальных сайтов рестрикции на NotI и BamHI между укороченными генами cryI в тандемных плазмидах pBD560 и pBD650 обусловлено последовательностями полилинкеров. Манипулирование ДНК и конструирование гибридного токсина. Все приемы манипулирования рекомбинантной ДНК проводили в соответствии с Sambrook et al.,1989 (в "Molecular Cloning, A Laboratory Manual: Cold Spring Harbour Press, Cold Spring Harbour), причем последовательность ДНК расшифровывали дидезокситрифосфатным методом с флуоресцентным окрашиванием дидезоксинуклеотидов, используя анализатор последовательности нуклеотидов Applied Biosystems 370A. Выявленная степень подобия cryI генов позволяет проводить внутримолекулярную рекомбинацию in vivo. В результате получают пару тандемных плазмид, каждая из которых несет два укороченных гена кристаллических белков, перекрывающиеся только в доменах II и III. Таким образом, рекомбинация происходит только в областях, кодирующих домены II и III. Рамочные рекомбинации, которые можно отбирать, используя рестриктазы для гидролиза, производят плазмиды, дающие возможность экспрессии полномерных гибридных протоксинов с молекулярной массой 140 кДа. Для получения in vivo рекомбинантов тандемную плазмиду (pBD560 или pBD650; фиг.2) переносят в JM101. ДНК в количестве 5 мкг выделяют из отдельно полученных рекомбинантов и гидролизуют с помощью NotI и BamHI, делая разрез между двумя укороченными cryI генами для селекции из нерекомбинантов, и ДНК трансформируется в E.coli XL1-голубую. Затем выращивают 5 отдельных колоний и анализируют характер белков и содержание плазмид. Гибридные токсины CryIC-CryIE и CryIE-CryIC получают с использованием соответственно тандемных плазмид pBD560 и pBD650, допуская их рекомбинацию в rесА+фон, выделяют ДНК, гидролизуют и переносят в штамм rесА, как описано выше. 100 колоний из 20 отдельных экспериментов анализируют электрофорезом в полиакриламидном геле с додецилсульфатом натрия. Из этих колоний 85% дают белок молекулярной массой 140 кДа, задаваемый рамочными рекомбинациями соответственно генов сrуIС и cryIE, cryIE и cryIC. E.coli продуцируют белки CryI в виде кристаллов, растворяющихся in vitro при высоких значениях рН. Приблизительно 15% гибридных токсинов, полученных описанным способом, растворяются при высоких значениях рН. Рекомбинанты, продуцирующие растворимые гибридные токсины, сначала классифицируют с использованием рестриктазы, затем определяют положение кроссовера в селекционированных гибридах каждого класса, анализируя последовательность ДНК. Все кроссоверы, которые приводят к получению растворимых гибридных токсинов, обнаруживаются очень близко от домена III или в нем. Очистка и анализ белков. Кристаллические белки выделяют в основном по методу Convents et al. (J. Biol. Chem. , 265, р.1369-1375; Eur. J. Biochem., 195, р.631-635). Вкратце, рекомбинантные E.coli выращивают при 30oС в 250 мл в ТВ-среде до ОП660 10-15. Кристаллы, выделенные из лизата E.coli, растворяют при инкубации в течение 2 часов в присутствии 20 мМоль Nа2СО3, 10 мМоль дитиотреитола, 100 мМоль NaCl при рН 10 и температуре 37oС. рН раствора понижают до 8 с помощью 100 мМоль буфера Tris-HCl, и инкубируют в присутствии трипсина. Раствор токсина диализируют против 20 мМоль буфера Tris-HCl и 100 мМоль NaCl при рН 9. Затем токсичный фрагмент очищают на колонке Mono Q 5/5, включенной в систему (FPLC) скоростной жидкостной хроматографии белков (Pharmacia LKB Biotechnology). Белки выделяют электрофорезом в полиакриламидном геле, содержащем 7,5% додецилсульфата натрия. Биохимический анализ и выделение токсичных фрагментов с молекулярной массой 65 кДа. Выделенные кристаллы очищенных белков CryIC, CryIE и гибридные белки растворяют при высоком значении рН и инкубируют с трипсином. Подобно белкам CryIC и СryIЕ все растворимые гибридные токсины при кроссовере в домене III превращаются в устойчивые фрагменты с молекулярной массой 65 кДа. Фрагменты с молекулярной массой 65 кДа можно очищать анионообменной хроматографией при таких же условиях, как и родительские белки. Tрипсин полностью разрушает гибриды F59 и F71, которые имеют кроссоверы в домене II. Несмотря на то, что эти гибриды не осаждаются в виде нерастворимых агрегаций, по-видимому, индуцированные трипсином места расщепления в родительских белках могут подвергнуться действию трипсина. Поэтому фрагменты с молекулярной массой 65 кДa не могут быть выделены из гибридов F59 и F71. В табл. 1 показана структура 5 гибридных токсинов CryIE-CryIC (G27, Н8, Н17, Н13, Н7) и 4 CryIC-CryIE гибридных токсинов (F59, F71, F26, Е7) в сравнении с белками CryIE и CryIC, из которых они получены. Аминокислотные последовательности гибридных токсинов CryIE-CryIC, содержащих токсичные фрагменты согласно изобретению, занимают положения до аминокислоты 1189 родительского белка CryIC, а гибридных токсинов CryIC-CryIE - до аминокислоты 1171 родительского белка CryIE. В табл. 1 приведены также значения относительной инсектицидной активности этих гибридных токсинов в сравнении с белками CryIE и CryIC. Большинство биологических анализов было проведено на очищенных фрагментах токсинов. Белки cryIC занимают положения от приблизительно аминокислоты 28 до 627, a cryIE - до 612. Полная длина последовательностей протоксинов указана в скобках. Определение токсичности для насекомых и инсектицидная активность cryICI cryIE продуктов гибридных генов. Бактериальные культуры концентрируют до ОП660 6,0 и 100 мкл наносят в виде точек на искусственную питательную среду на матрас для культуры тканей, имеющий 24 лунки. Наряду с этим разбавленные образцы очищенных токсинов наносят на питательную среду. Личинки во втором инстаре насекомых S.exigua, M. bras-sicae или M. sexta содержат на этой питательной среде (16-кратное разбавление на образец) 5 дней, после чего учитывают массу личинок. Относительный рост (равновесную концентрацию, снижающую рост на 50%) определяют по отношению средней массы личинок, содержащихся на питательной среде с токсином, к средней массе контрольных личинок, содержащихся на питательной среде без токсина. Насекомых вида M.sexta для кладки яиц поставляет Carolina Biological Supply Company, North Carolina, US. Токсичные фрагменты, кодированные продуктами гибридных генов, испытывают на активность против трех различных видов насекомых, описанных выше. Насекомые вида M. sexta восприимчивы как к CryIC, так и СrуIЕ. По их чувтствительности к трипсину можно предположить, что гибриды F59 и F71 не обладают активностью против этого насекомого (табл. 1). Несмотря на то, что Н7 конвертируется трипсином в устойчивые белки с молекулярной массой 65 кДа, он не токсичен для M. sexta. Все остальные гибриды, представленные в табл. 1, токсичны и, по-видимому, имеют нативную биологически активную конформацию. Фрагмент белка CrylC молекулярной массой 65 кДа в отличие от белка CryIE высокотоксичен по отношению к видам S. exigua и M.brassicae. Токсин G27 (табл. 1; фиг. 2), гибрид CryIE-CryIC с кроссовером в месте соединения доменов II и III активен по отношению к обоим видам насекомых. Это указывает на то, что домен III белка CryIC придает ему полную активность по отношению к S.exigua и M.brassicae. Гибрид Н8, который отличается от G27 только тремя аминокислотными остатками (см. фиг.3), будучи активным против M.sexta, не активен против S.exigua и M.brassicae. Антипод гибрида G27 - гибрид F26 (табл. 1, фиг.3), в котором домен III белка CryIC заменен доменом III белка CryIE, не активен против S.exigua или M. brassicae. Несмотря на то, что он токсичен по отношению к M.sexta, последовательности белка CryIC, включающие положения от аминокислот 28 до 462, вероятно, не достаточны для поражения S.exigua и M.brassicae. Последовательности белка CryIC только тогда восстановливают инсектицидную активность против этих насекомых, когда в этом гибриде (Е7) присутствуют аминокислотные остатки не ниже положения 602. В этом описании показано, что аминокислотные остатки в положениях 478-602 последовательности белка CrylC могут придать белку CrylE высокую инсектицидную активность против S.exigua и M.brassicae. Биотинилирование кристаллических белков и их анализ по степени связывания. Биотинилирование проводят с использованием биотин-N-гидроксисукцинимиидного эфира по существу таким образом, как оно описано изготовителем (Amersham). 1 мг кристаллического белка инкубируют в течение одного часа с 40 мкл биотинилирующего реагента в присутствии 50 мМоль NаНСО3, 15 мМоль NaCl при рН 8 и температуре 20oС. Раствор заливают в колонку, заполненную Sephadex 25, уравновешенным тем же буфером, содержащим 0,1% бычьего сывороточного альбумина для удаления несвязанного биотина, и образцы фракций наносят пятнами на нитроцеллюлозную мембрану. Фракции, содержащие биотинилированные кристаллические белки, визуализируют с помощью конъюгата стрептавидин-пероксидаза (Amersham), который катализирует окисление люминола, вызывающего хемилюминесценцию (ECL, Amersham), и сливают. Мембранные везикулы собирают по Wolfersberger et al. (Сотр. Biochem. Physiol., 86a, p.301-308, 1987) и, кроме того, еще раз промывают изолирующим буфером, содержащим 0,1% среды Tween-20. Связывание биотинилированных кристаллических белков с мембранными везикулами (100 мкг/мл) проводят в 100 мкл среды PBS, содержащей 1% бычьего сывороточного альбумина и 0,1% среды Tween-20 при рН7,6. Везикулы (20 мкг белка) инкубируют с 10 мг биотинилированных кристаллических белков в течение одного часа при 20oС в присутствии или при отсутствии 1000-кратного избытка немеченых кристаллических белков. Затем везикулы снова отделяют центрифугированием в течение 10 мин при 14000 g в центрифуге Eppendorf, дважды промывают связующим буфером, снова суспендируют в образцовом буфере, денатурируют нагреванием и вводят в 7,5%-ный полиакриламидный гель. После электрофореза белки блотируют на нитроцеллюлозные мембраны и биотинилированные кристаллические белки, которые повторно выделены с везикулами, визуализуют, инкубируя нитроцеллюлозу с конъюгатом стрептавидин-пероксидаза (Amersham), который катализирует окисление люминола, вызывающего хемилюминесценцию (ECL, Amersham). Поскольку связывание с эпителиальными клетками средней кишки - основная часть механизма действия кристаллических белков, их связывание мембранными везикулами S.exigna исследуют в экспериментах на гетерологическую конкуренцию. Конкурентные эксперименты показывают, что связывание меченого белка CryIC (фиг. 4А, цепочка а) и меченого гибрида F26 (не показан) может быть исключено при избытке немеченого белка CryIC (цепочка b) или гибрида F26 (цепочка е), но не при избытке гибрида G27 (цепочка с) или белка CryIE (цепочка d). Кроме того, связывание меченого гибрида G27 (фиг.4В, цепочка а) и меченого белка CryIE (не показан) может быть исключено при избытке гибрида G27 (цепочка b) или белка CryIE (цепочка d), но не при избытке белка CryIC (цепочка а) или гибрида F26 (цепочка е). Это позволяет сделать вывод, что G27 и CryIE "узнают" те же сайты связывания на мембранах средней кишки S. exigna и что эти сайты отличаются от сайтов, "узнаваемых" белком СrуIС и гибридом F26. Совместные испытания на токсичность и степень связывания показывают, что G27 обладает такой же токсичностью, как и CryIC, но связывается с иным рецептором. Поскольку насекомые могут приобретать сопротивляемость к кристаллическим белкам, изменяя показатели связывания рецепторов, G27 может быть полезен для контроля сопротивляемости насекомых в качестве варианта замены CryIC. Экспрессия генов гибридного токсина cryIE/cryIC в гетерологичных системах. Ген cryIE/cryIC гибридного токсина G27 экспрессируется в E.coli, и генный продукт проявляет по меньшей мере такую же инсектицидную активность (по меньшей мере против Spodoptera) как CryIC. Более того, этот продукт проявляет повышенную активность, если ген экспрессируется в штамме Bacillys thuringienses (см. ниже). Ген, кодирующий гибридный токсин G27, вводят в подходящий вектор-челнок, который затем вводят в соответствующего хозяина B.t. Такие трансформированные клетки культивируют, а получаемый от обеих цельных культур токсин и очищенные кристаллы проверяют на инсектицидную активность. Конструирование вектора-челнока, содержащего G27, трансформация штамма Bt 51 и очистка от него токсичного белка. Ген, кодирующий гибрид G27 (3400 пар оснований), отщепляют от плазмиды, индуцирующей экспрессию рКК233 Е.соli, с использованием Nсо1 и Хhо1. Сайт Xho1 заполняют фрагментом Кленова, т.е. фрагментом ДНК-полимеразы I от Е. соli. Полученный фрагмент лигируют с Nco1/Sma1, гидролизованной рSB635 (pBluescript KS+, PcryIC и CryIA(c) терминаторы транскрипции). Полученную плазмиду рSВ454 гидролизуют с Ара1 и Nоt1, образуя фрагмент с 4200 пар оснований, несущий промотор, гибрид G27 и терминатор. Этот фрагмент лигируют с Ара1/Not1 гидролизованной рSВ634 (челнок-вектор, содержащий рВС16.1 и pBluescript KS+), образуя рSВ456 (см. фиг.5). Плазмиду ДНК, выделенную из E.coli, ДН10В, используют для трансформации B.t.51, штамма B.t., не вырабатывающего кристаллический токсин. Положительные изоляты устойчивы к тетрациклину, что говорит о присутствии pSB456, и содержат большие включения, соответствующие белку с молекулярной массой 135 кДа (выявлено с помощью электрофореза в полиакриламидном геле с додецилсульфатом натрия). Образцы гибридного токсина G27 готовят из культур трансформированного штамма Bt51, выращенного спорообразованием при 30oС в среде цистеин-тетрациклин10. Биологический анализ инсектицидности (табл. 2) проводят как на полных цельных культурах, так и на отмытых препаратах кристаллического белка. Контрольные образцы включают Bt51 (рSВ440), содержащий токсин CryIC, и Bt51 (рSВ636), содержащий CryIE. Концентрацию токсина определяют электрофорезом в полиакриламидном геле с додецилсульфатом натрия. Несмотря на то, что изобретение описано на примере получения гибридного токсина G27, специалисту будет понятно, что, пользуясь информацией из этого описания, можно получить многие другие гибридные токсины с улучшенными инсектицидными показателями. Например, в SEQ ID No.7 и 8 представлены другие гибридные токсины согласно изобретению, которые обладают повышенной инсектицидной активностью. Можно получить гибридные токсины, содержащие домен III белка CryIC и N-конец любого иного белка Cry. Что касается биологических испытаний, триансформанты, несущие гибридный токсин, можно вырастить в SOP средах для ускорения испытаний и уменьшения объемов используемого материала. Кроме того, ген, кодирующий G27 и/или другие гибридные токсины, согласно изобретению можно перенести в кодирующие токсин штаммы B.t. и/или интегрировать в хромосому выведенных штамов B.t. или даже ввести в геномы растений для обеспечения инсектицидной активности in situ в этом растении. В этом случае особенно предпочтительно рекомбинантную ДНК, кодирующую токсины, модифицировать так, чтобы кодоны, предпочтительные для растения, которому эта рекомбинантная ДНК предназначена, были использованы так, что экспрессией такой модифицированной ДНК в указанном растении синтезируется белок, по существу сходный с белком, синтезируемым при экспрессии немодифицированной рекомбинантной ДНК в организме, для которого белковые компоненты гибридного токсина или фрагментов токсина являются эндогенными.
ФОРМУЛА ИЗОБРЕТЕНИЯ1. Фрагмент гибридного токсина Bacillus thuringiensis (B. t), обладающий инсектицидной активностью, где ДНК гена, кодирующего указанный фрагмент, получен посредством единичного случая рекомбинации, содержащего на своем С-терминальном конце домен III белка СrуС или СrуА, при условии, что N-терминальная область фрагмента содержит домен II белка, выбранного из группы, включающей CryIA(a), CryIA(b), CryIA(c), CryIB, CryIC, CryID, CryIE, CryIF, CryIG, CryIH, CryIIA, CryIIB, CryIIC, CryIIIA, CryIIIB, CryIIIB(b), CryIVA, CryIVB, CryIVC, CryIVD, CYTA, CryXI(IIIC), CryX2(IIID), СrуХ3, CryV и CryX4, и этот фрагмент токсина B. t. связывается с рецептором, расположенным в кишке насекомого, отличающимся от рецептора, который связывается с CryC- или СrуА- белком. 2. Фрагмент гибридного токсина B. t. по п. 1, отличающийся тем, что он не связывается с рецептором, связывающимся с CryIC. 3. Фрагмент токсина по любому предшествующему пункту, отличающийся тем, что N-терминальная область содержит домен II белков CryIE, CryIB, CryID или CryIA(a), CryIA(b), CryIA(c) и домен III белка CryIC. 4. Фрагмент токсина по любому предшествующему пункту, отличающийся тем, что N-терминальная область кроме домена II, содержит домены I белков CrylE, CryIB, CryID, CryIA(a), CryIA(b) или CryIA(c), и домен III белка CryIC. 5. Фрагмент токсина по любому предшествующему пункту, отличающийся тем, что он имеет на своем С-терминальном конце область, содержащую последовательность от аминокислотного положения 454 до положения 602 белка СrуIC. 6. Фрагмент токсина по любому предшествующему пункту, отличающийся тем, что он имеет на своем С-терминальном конце область, содержащую последовательность от аминокислоты в положении 478 до аминокислоты в положении 602 белка CryIC. 7. Фрагмент токсина по п. 1, отличающийся тем, что он содержит последовательность аминокислот приблизительно от аминокислоты 1 до приблизительно аминокислоты 620 в SEQ ID No. 6 или последовательность аминокислот от приблизительно аминокислоты 1 до приблизительно аминокислоты 620 в SEQ ID No. 6, в которой по отношению к указанной последовательности произведена по меньшей мере одна из следующих замен: Ilе в положении 609 заменен Leu; Ala в положении 618 заменен Glu; Ser в положении 620 заменен Туr. 8. Фрагмент токсина по п. 1, отличающийся тем, что он содержит последовательность аминокислот приблизительно от аминокислоты 1 до приблизительно аминокислоты 627 в SEQ ID No. 8 или последовательность аминокислот от приблизительно аминокислоты 1 до приблизительно аминокислоты 627 в SEQ ID No. 8, в которой по отношению к указанной последовательности произведена по меньшей мере одна из следующих замен: Ilе в положении 617 заменен Leu; Ala в положении 625 заменен Glu; Ser в положении 627 заменен Туr. 9. Рекомбинантная ДНК, кодирующая фрагмент гибридного токсина Bacillus thuringiensis по любому из пп. 1-8, содержит фрагмент нуклеотидной последовательности от нуклеотида 1 до приблизительно нуклеотида 1860, приведенной в SEQ ID No. 5, или фрагмент нуклеотидной последовательности от нуклеотида 1 до приблизительно нуклеотида 1881, приведенной в SEQ ID No. 7. 10. Рекомбинантная ДНК по п. 9, отличающаяся тем, что она модифицирована так, что из нее исключены известные основные элементы нестабильности мРНК или сигналы полиаденилирования, и/или ее кодоны являются предпочтительными для организма, которому эта рекомбинантная ДНК предназначена, и ее используют так, что при экспрессии такой модифицированной ДНК в указанном организме синтезируется белок, по существу, сходный с белком, синтезируемым при экспрессии немодифицированной рекомбинантной ДНК в организме, содержащем эндогенные компоненты белка гибридного токсина или фрагментов токсина. 11. Рекомбинантная ДНК по любому из пп. 9 и 10, которая содержится в векторе. 12. Способ борьбы с насекомыми, который заключается в обработке их фрагментом гибридного токсина по любому из пп. 1-8 или композициями, содержащими этот токсин.Популярные патенты: 2007081 Способ биологической борьбы с вредителями капусты ... способу и предложенному при расстоянии посеянных нектароносов от капустного поля около 60 м. Как видно из табл. 2 и 3, эффективность влияния энтомофагов на вредителей капусты в предложенном способе выше в среднем на 27% , а в отношении трихограммы, птеромалуса и апантелеса - на 30-45% . Это можно объяснить более ранним началом цветения рекомендуемой смеси нектароносов в предложенном способе, а значит и доступностью нектара энтомофагам, многие из которых появляются на 2-3 недели раньше их основных хозяев - вредителей капусты. Поэтому наличие в этот период нектара увеличивает срок жизни и плодовитость энтомофагов в 1,5-2 раза, что помогает им дождаться появления подходящей для ... 2065260 Гидравлическая система самоходной сельскохозяйственной машины ... которые управляются посредством соответственных гидроцилиндров 7, 8 и 9. Привод ведущих колес машины включает насос 10, реверсивный гидромотор 11, тягу 12 и педаль включения 13. На раме 1 установлен также секционный гидрораспределитель 14, а под педалью 13 установлена также блокировочный сервоцилиндр 15, соединенный магистралями 16 и 17 (см. фиг. 2) с блоком 18 последовательно соединенных управляемых обратных клапанов (УОК) 19, 20, 21 и блоком 22 параллельно соединенных клапанов 23, 24, 25. В управляемые магистрали 26, 27, 28 УОК 19, 20, 21 включены параллельно соединенные и противоположно направленные редукционные клапаны (РК) 29, 30, 31 и обратные клапаны (ОК) 32, 33, 34. В свою ... 2088063 Широкозахватный сельскохозяйственный агрегат ... бруса и с корпусами гидроцилиндров свода-развода задних звеньев, штоки которых шарнирно соединены с задними звеньями, гидроцилиндры поворота задних звеньев вокруг своих осей корпусами шарнирно соединены с задними звеньями, а штоками - с вертикальными шарнирами звеньев, при этом гидроцилиндры свода-развода снабжены гидрозапорным блоком. Такое выполнение механизма перевода агрегата в рабочее и транспортное положение позволяет при отключении гидроцилиндров свода-развода задних звеньев после окончания перевода агрегата в рабочее или транспортное положение свободно перетекать рабочей жидкости между полостями гидроцилиндров, обеспечивая тем самым копирование рельефа поля каждым ... 2056755 Способ регулирования роста овощных культур ... отбирали среднюю пробу и определяли содержание нитратов в листьях растений салата. Повторность опыта пятикратная. Данные представлены в табл. 8 Данные, приведенные в табл. 8, указывают на то, что предлагаемый способ значительно (на 30%) эффективнее способа-прототипа. Кроме того, как видно из табл. 7, эффект препарата Сапфир основан на не прямом физиологическом действии зеатина и зеатинрибозида, а на синергическом воздействии этих регуляторов и других коэкстрактивных веществ, содержащихся в цитокининовом препарате, о чем так же свидетельствует отсутствие подобного эффекта у других известных цитокининов кинетина и полистимулина К (табл. 8). Таким образом, представленные в табл. 1-8 ... 2288561 Устройство для предпосевной обработки семян растений ... семян и уменьшить утечки электромагнитной энергии, но при этом происходит обработка только потоком электромагнитной энергии, не изменяется влажность семян и не оказывается фунгистатического или бактерицидного воздействия на семена.Известно устройство для обработки семян холодной плазмой (см. патент США №6543460, МПК В 08 В 7/04, опубликован 08.04.2003), включающее горизонтально установленную с возможностью вращения цилиндрическую реакционную камеру из пирекса, закрытую с торцов дисками из нержавеющей стали. Коаксиально барабану установлено два полуцилиндрических электрода из меди. Верхний электрод соединен с высокочастотным генератором, а нижний электрод заземлен. ... |
Еще из этого раздела: 2274986 Способ посева семян трав и кустарников для создания пастбищ на опустыненных землях и почвообрабатывающее орудие для его осуществления 2087614 Способ создания травяного газонного покрытия открытых спортивных площадок и ухода за ним 2165141 Тепличный гидропонный комплекс 2210910 Способ обработки растений и используемая в нём композиция для защиты растений 2233582 Устройство для охлаждения молока 2471341 Стойло, устройство в стойле и способ монтажа указанного устройства 2091380 Производные пиколиновой кислоты или их кислотно-аддитивные соли, способ их получения, нербицидная композиция и способ борьбы с сорняками 2172085 Способ управления групповым вождением машин 2302109 Способ снижения уровня никеля и свинца в крови и молоке коров техногенной провинции 2175189 Способ регенерации растений сорго в культуре in vitro |
Изобретения в сельском хозяйстве
Обработка почвы в сельском и лесном хозяйствах
Посадка, посев, удобрение
Уборка урожая, жатва
Обработка и хранение продуктов полеводства и садоводства
Садоводство, разведение овощей, цветов, риса, фруктов, винограда, лесное хозяйство
Новые виды растений или способы их выращивания
Производство молочных продуктов
Животноводство, разведение и содержание птицы, рыбы, насекомых, рыбоводство, рыболовство
Поимка, отлов или отпугивание животных
Консервирование туш животных, или растений или их частей
Биоцидная, репеллентная, аттрактантная или регулирующая рост растений активность химических соединений или препаратов
Хлебопекарные печи, машины и прочее оборудование для хлебопечения
Машины или оборудование для приготовления или обработки теста
Обработка муки или теста для выпечки, способы выпечки, мучные изделия
|
|
||