Способ получения гибридов между культурным картофелем solanum tuberosum и 1евn диплоидными дикими видами картофеляПатент на изобретение №: 2505957 Автор: Ермишин Александр Петрович (BY), Воронкова Елена Васильевна (BY), Полюхович Юлия Владимировна (BY), Маханько Ольга Владимировна (BY) Патентообладатель: Государственное научное учреждение "Институт генетики и цитологии Национальной академии наук Беларуси" (BY) Дата публикации: 10 Февраля, 2014 Начало действия патента: 13 Августа, 2012 Адрес для переписки: 220072, Республика Беларусь, г.Минск, ул. Академическая, 27, ГНУ "Институт генетики и цитологии Национальной академии наук Беларуси", ОНИР, А.П. Быченко Изображения    Изобретение относится к селекции растений картофеля Solanum tuberosum. Изобретение представляет собой способ, включающий гибридизацию между 1EBN диплоидными дикими видами картофеля и видом (линией)-посредником и последущее беккроссирование полученных гибридов дигаплоидами Solanum tuberosum, где в качестве материнских форм при гибридизации между 1EBN диплоидными дикими видами картофеля и видом (линией)-посредником используют растения, отобранные в поколении F2 гибридов между дигаплоидами Solanum tuberosum и диким диплоидным видом картофеля Solanum verrucosum, несущие в гомозиготном состоянии S гены, привнесенные от Solanum verrucosum. Данное изобретение позволяет повысить выход гибридных семян, фертильность межвидовых гибридов, их способность к клубнеобразованию и способность к гибридизации с культурным картофелем. 5 ил. Изобретение относится к селекции растений, например, картофеля Solanum tuberosum, и может использоваться в сельском хозяйстве. В селекции растений, например, картофеля Solarium tuberosum, применяют метод отдаленной гибридизации между культурным картофелем и 1EBN диплоидными дикими видами картофеля, например, Solanum bulbocastanum, Solanum pinnatisectum, Solarium polyadenium, Solarium commersonii и др. с целью интрогрессии в селекционный материал ценных генов диких видов. 1EBN диплоидные дикие виды картофеля имеют диплоидный набор хромосом (2n=2x=24), в отличие от культурного картофеля, который является автотетраплоидом (2n=4x=48, 4EBN). Поэтому для вовлечения их в гибридизацию с культурным картофелем целесообразно использовать дигаплоиды Solarium tuberosum (2n=2x=24, 2EBN). Однако гибридизация между этими видами и дигаплоидами Solarium tuberosum сильно затруднена из-за жестких межвидовых пре- и постзиготных репродуктивных барьеров. Известен способ получения гибридов между культурным картофелем Solarium tuberosum и 1EBN дикими видами картофеля, основанный на проведении промежуточных скрещиваний ("bridge crosses") для преодоления межвидовых репродуктивных барьеров [1]). Считается, что наряду с двумя несовместимыми видами, например диким видом и культурным картофелем, может быть третий вид картофеля (вид-посредник - "bridge species"), который скрещивается с диким видом, а полученные гибриды можно непосредственно, или после изменения их плоидности, или через гибридизацию с очередным видом-посредником вовлечь в гибридизацию с культурным картофелем, что в итоге позволяет осуществить интрогрессию ценных генов дикого вида в селекционный материал. В качестве посредника для вовлечения в скрещивания дикого диплоидного вида картофеля S.bulbocastanum (2x, 1EBN) с S.tuberosum в этом способе использовали аллотетраплоидный (4х, 2EBN) вид S.acaule. У полученных триплоидных двухвидовых гибридов АВ (acaule-bulVocastanum) удвоили число хромосом, затем гексаплоиды S.acaule × S.bulbocastanum 6x (АВ)2 скрестили с видом-посредником S.phureja (2x, 2EBN), получив тетраплоидный гибрид 4х (АВ)2P, который можно было скрещивать с сортами культурного картофеля S.tuberosum (4x, 4EBN). Недостатки способа следующие. Из-за низкой эффективности гибридизации на всех этапах его осуществления требуется проведение большого объема скрещиваний. Длинная цепочка скрещиваний с посредниками и манипуляции с плоидностью гибридов занимают длительный период времени, удвоение плоидности гибридов связано с использованием сложных методов митотического удвоения хромосом. В результате эффективность метода оказалась невысокой: было получено 36 четырехвидовых гибридных растении АВРТ от опыления, в общей сложности, 18 616 цветков. Наиболее близким техническим решением является способ получения гибридов между культурным картофелем S.tuberosum и 1EBN дикими видами картофеля, который основан на получении промежуточных гибридов с участием дикого диплоидного вида S.verrucosum (2n=2x=24, 2EBN), характерной особенностью которого является отсутствие пестичных S-РНКаз [2]. Способ включает гибридизацию между образцами дикого диплоидного вида S.verrucosum, которые используют в качестве материнских форм, и мужски фертильными образцами 1EBN диких видов картофеля, которые используют в качестве опылителей. Полученные промежуточные гибриды беккроссируют дигаплоидами S.tuberosum или диплоидными межвидовыми гибридами, а затем дигаплоидами S.tuberosum. Для переноса интрогрессированных от 1EBN диких видов картофеля ценных генов в селекционный материал S. tuberosum (2n=4x=48) отобранные в беккроссных поколениях генотипы переводят на тетраплоидный уровень с помощью митотического или мейотического удвоения хромосом. Отсутствие пестичных S-РНКаз у S.verrucosum позволяет решить проблему презиготной несовместимости, имеющей место между многими 1EBN дикими видами и 2EBN видами картофеля: пыльца любого вида картофеля, в том числе 1EBN диких видов, способна беспрепятственно расти в пестиках S.verrucosum. Однако образование жизнеспособных гибридных семян затруднено из-за нарушения развития их эндосперма, обусловленного различиями EBN родительских видов. Тем не менее, можно предположить, что благодаря отсутствию презиготных барьеров в скрещиваниях между S.verrucosum (2EBN) и 1EBN видами возможно завязывание небольшого количества семян вопреки правилу совпадения эффективной плоидности родительских видов. Этот способ имеет следующие недостатки. Завязываемость жизнеспособных семян при гибридизации между видом-посредником S.verrncosum и некоторыми 1EBN дикими видами картофеля, как правило, очень низкая. Так, с помощью описанного способа не были получены гибриды между S.verrucosum и одним из наиболее ценных для селекции диких видов картофеля S.bulbocastanum. He удалось получить такие гибриды и Dinu [3], несмотря на большой объем скрещиваний и использование специальных приемов (культура гибридных зародышей in vitro). Полученные гибриды между S.verrucosum и 1EBN дикими видами картофеля практически не образуют клубни в условиях Беларуси, что затрудняет их беккросснрование дигаплоидами S.tuberosiim. Эти гибриды с трудом скрещиваются с дигаплоидами S.tuberosum при использовании в качестве материнских форм из-за действия факторов презиготной несовместимости, полученных от 1EBN дикого вида (пестичных РНКаз 1EBN дикого вида). Мужски фертильные межвидовые гибриды не скрещиваются с дигаплоидами S.tuberosum при использовании в качестве опылителей по причине односторонней несовместимости (для успешной гибридизации требуются акцепторные формы S.tuberosum). Если их удается беккроссировать дигаплоидами S.tuberosum, используя в качестве материнских форм, получается мужски стерильное потомство, что является результатом взаимодействия цитоплазмы S.verrucosum с ядерными генами S.tuberosum (ядерно-цитоплазматическая мужская стерильность). Это не позволяет использовать мейотическое удвоение хромосом для перевода беккроссов на тетраплоидный уровень (не образуется фертильная нередуцированная пыльца). Сущность изобретения состоит в решении задачи повышения эффективности гибридизации между посредником и 1EBN дикими видами картофеля и упрощения процедуры беккроссирования полученных гибридов дигаплоидами и S.tuberosum. Решение задачи состоит в том, что в известном способе, включающем получение промежуточных гибридов между 1EBN диплоидными дикими видами картофеля и видом (линией)-посредником, не образующим пестичных РНК-аз, и последующее беккроссирование полученных гибридов дигаплоидами S.tuberosum, с целью повышения выхода гибридных семян, способности промежуточных гибридов к клубнеобразованию, получения мужски фертильного поколения беккроссов получают и используют в качестве материнских форм при гибридизации с 1EBN дикими видами растения, отобранные в поколении F2 гибридов между дигаплоидами S.tuberosum и диким диплоидным видом картофеля S.verrncosum, несущие в гомозиготном состоянии S гены, привнесенные от S.verrucosum (далее - SvSv-линии). У картофеля презиготные барьеры ассоциируют с функционированием так называемого S-локуса (локализован на хромосоме I). Результат гибридизации как внутривидовой, так и межвидовой, определяется взаимодействием продуктов пестичного и пыльцевого компонентов S-локуса. Продуктами пестичного компонента S-локуса являются S-РНКазы, которые содержатся в пестиках вне зависимости от факта опыления, могут свободно проникать в цитоплазму пыльцевых трубок и, действуя как ингибитор процесса трансляции, останавливать их рост. Этот своеобразный барьер могут преодолеть лишь те пыльцевые трубки, которые содержат определенные продукты, способные дезактивировать соответствующие РНКазы. В качестве кандидатов в продукты пыльцевого компонента S-локуса рассматривают так называемые SLF-протеины (F-box proteins of S locus), которые могут узнавать соответствующие S-РНКазы и запускать систему деградации S-РНКаз. Есть основания полагать, что межвидовая несовместимость у картофеля обусловлена тем, что процесс взаимодействия пыльцевого и пестичного компонентов S-локуса при гибридизации отдаленных в эволюционном отношении видов нарушен, поскольку дезактивация соответствующих пестичных РНКаз может произойти только с помощью SLF-протеинов того же или близкородственного вида. 1 EBN диплоидные виды картофеля, многие из которых произрастают в Мексике, - группа наиболее древних с точки зрения эволюции клубненосных видов Solanum. Неслучайно поэтому, что для многих 1 EBN диплоидных видов картофеля характерна высокая степень выраженности презиготных барьеров несовместимости при гибридизации их с дигаплоидами S.tuberosum. Особенно это относится к ценному для селекции виду картофеля S.bulbocastanum, при гибридизации которого с днгаплоидами S.tuberosum имеет место полное ингибирование пыльцы в пестиках вне зависимости от использования разных генотипов компонентов скрещивания [4]. Постзиготными считаются барьеры, возникающие после проникновения пыльцевых трубок в завязь и оплодотворения. Прежде всего, они обусловлены гибелью зародышей вследствие недоразвития эндосперма и связаны с понятием EBN (балансовое число эндосперма) вида. Каждому виду картофеля предписывается определенный специфический показатель (его EBN), значение которого определяет поведение этого вида (формирование эндосперма гибридных семян) при гибридизации с другими видами. Для успешной гибридизации EBN родительских форм должны совпадать, чтобы эндосперм, а следовательно, и семя развивались нормально. EBN вида может быть не равен его плоидности, оно является мерилом его "эффективной плоидности". Для каждого вида определена его эффективная плоидность, что позволило сгруппировать виды по группам скрещивания - 2х (1EBN), 2x (2EBN), 3х (2EBN), 4х (2EBN), 4х (4EBN) и 6х (4EBN) [5]. Благодаря наличию в гомозиготном состоянии S генов (Sv Sv), привнесенных от S.verrucosum, SvS v-линии не образуют пестичных РНКаз. Поэтому они, так же как и S.verrucosum не имеют презиготных барьеров в скрещиваниях с 1EBN дикими видами картофеля при использовании их в качестве материнских форм. Соответственно, в таких скрещиваниях возможно завязывание небольшого количества гибридных семян вопреки правилу совпадения эффективной плоидности родительских форм. По сравнению с S.verrucosum растения SvSv-линий отличаются культурным типом куста, большей мощностью, цветут более интенсивно и продолжительно, способны завязывать больше гибридных семян, способны формировать клубни в условиях Беларуси. Промежуточные гибриды между SvSv-линиями и 1EBN дикими видами картофеля в качестве одного из родителей (SvSv-линии) имеют относительно культурную форму. Благодаря этому они лучше приспособлены к климатическим условиям Беларуси (умеренные широты, длинный летний день), в силу чего обладают более высокой способностью к клубнеобразованию. Они несут большее количество признаков культурного картофеля, и меньшее количество нежелательных для селекции признаков дикого вида, чем растения S.verrucosum. Поэтому требуется меньше поколений беккросса дигаплоидами S.tuberosum промежуточных гибридов между 1EBN дикими видами картофеля и посредником, чтобы усилить культурные признаки и избавиться от нежелательных признаков дикого вида. Это свойство SvSv-линий может быть усилено в ходе последующей селекции, включающей их беккроссирование дигаплоидами S.tuberosum в сочетании с отбором гомозигот SvS v. Гибриды между SvSv-линиями и 1EBN дикими видами картофеля более мощные, чем гибриды между S.verrucosum и 1EBN дикими видами картофеля. Они цветут более обильно и в течение более длительного времени, что упрощает процедуру их беккроссирования дигаплоидами S.tuberosum. Растения промежуточных гибридов между посредником (SvS v-линией или S.verrucosum) имеют следующее строение S-локуса: SvSд.в, где Sv - аллель S-гена от Solanum verrucosum, а Sд.в. - аллель S-гена от 1EBN дикого вида, использованного в качестве опылителя. Соответственно, в пестиках таких гибридов будет образовываться S-РНКаза дикого вида (аллель Sv от S.verrucosum является нефункционирующим). Следует ожидать, что в силу гемизиготности активность пестичной РНКазы у гибридов будет ниже, чем у родительского дикого вида, поэтому презиготные барьеры при беккроссировании гибридов дигаплоидами S.tuberosum будут выражены не так сильно, как при гибридизации непосредственно между дигаплоидами S.tuberosum и 1EBN дикими видами картофеля. В результате можно рассчитывать на получение небольшого количества семян первого поколения беккросса (ВС1), даже при использовании промежуточных гибридов в качестве материнских форм. Получение последующих поколений беккросса культурным картофелем значительно упрощается в силу того, что среди ВС1 резко уменьшается доля генотипов с аллелем Sд.в, то есть образующих S-РНКазы дикого вида. Однако промежуточные гибриды и их беккроссное потомство на основе SvSv -линий имеют цитоплазму S.tuberosum, что обеспечивает их мужскую фертильность. В то же время практически все гибриды между S.verrucosum и дигаплоидами S.tuberosum мужски стерильны (ядерно-цитоплазматическая мужская стерильность) [6]. Мужская фертильность промежуточных гибридов и их беккроссного поколения благодаря наличию у них цитоплазмы S.tuberosum делает возможным использование мейотического удвоения хромосом у беккроссов для перевода их на тетраплоидный уровень с помощью мейотического удвоения хромосом, которое считается более простым и эффективным по сравнению с митотическим удвоением хромосом. Присутствие в пестиках промежуточных гибридов S-РНКазы дикого родительского вида может существенно снизить завязываемость семян при их беккроссировании дигаплоидами S.tuberosum, поскольку S-локус последних не кодирует образование ингибиторов РНКаз, способных дезактивировать S-РНКазы дикого родительского вида. Возможность использовать промежуточные гибриды в качестве опылителей позволяет решить проблему презиготной несовместимости с дигаплоидами S.tuberosum, поскольку в пыльце гибридов в силу расщепления по S-локусу будут пыльцевые зерна, несущие как аллель Sд.в., так и Sv. Пыльцевые зерна с аллелем Sv, как известно, кодируют ингибиторы S-РНКаз, способные дезактивировать S-РНКазы, находящиеся в пестиках дигаплоидов S.tiiberosum, относящихся к акцепторным генотипам по отношению к S. verrucosum (способным опыляться S.verrucosum) [7]. Однако в распоряжении исследователей не всегда имеются акцепторные генотипы S.tuberosum. Проблему беккроссирования промежуточных гибридов дигаплоидами S.tuberosum можно легко решить, если опылить пыльцой фертильных промежуточных гибридов материнские формы Sv Sv-линий. Половина потомства от таких скрещиваний будет гомозиготами SvSv, которые в силу отсутствия у них в пестиках S-РНКаз будут гарантированно скрещиваться с дигаплоидами S.tuberosum. Тот же результат достигается при опылении материнских форм S.verrucosum фертильной пыльцой промежуточных гибридов. Однако в этом случае потомство будет иметь значительно большую долю генома, полученную от диких видов, в связи с чем потребуется больше беккроссов культурным картофелем по сравнению с вариантом использования SvSv-линий. Кроме того, беккроссное потомство на цитоплазме S.verrucosum будет мужски стерильным. Сопоставительный анализ заявляемого решения с прототипом показывает, что заявляемый способ отличается от известного тем, что используют в качестве посредника вместо образцов S.verrucosum специально созданные SvS v-линии. Это дает возможность повысить, по сравнению с прототипом, эффективность гибридизации между посредником и 1EBN дикими видами картофеля. Более высокая жизнеспособность, интенсивность цветения и способность к клубнеобразованию промежуточных гибридов между посредником и 1EBN дикими видами картофеля позволяет упростить процедуру и уменьшить продолжительность их беккроссирования дигаплоидами S.tuberosum. Использование SvSv-линий в качестве материнских форм при опылении их фертильной пыльцой промежуточных гибридов позволяет получать мужски фертильное потомство, которое легко скрещивается с дигаплоидами S.tuberosum. Это значительно упрощает процедуру беккроссирования промежуточных гибридов дигаплоидами S.tuberosum и делает возможным использование мейотического удвоения хромосом у беккроссов для перевода их на тетраплоидный уровень. Фигура 1 представляет схему создания Sv Sv-линий. Фигура 2 представляет результаты гибридизации между рядом образцов S.verrucosum и 1EBN дикими видами картофеля. Фигура 3 представляет результаты гибридизации между рядом SvSv-линий и 1 EBN дикими видами картофеля, из которой видно, что завязываемость гибридных семян при использовании в качестве материнских форм SvSv-линнй выше, чем при использовании S.verrucosum. Фигура 4 представляет результаты оценки всхожести семян, клубнеобразования и фертильности пыльцы у гибридов F1 между S.verrucosum и 1EBN дикими видами картофеля и гибридов F1 между SvSv -линиями и 1EBN дикими видами картофеля. Эти результаты свидетельствует о более высокой по сравнению с прототипом всхожести семян, способности к клубнеобразованию и массе клубней с растения гибридов F 1 между SvSv-линиями и 1EBN дикими видами картофеля. Среди этих гибридов достоверно меньше стерильных и низкофертильных генотипов и больше генотипов со средней и высокой функциональной фертильностью пыльцы. Фигура 5 представляет результаты беккроссирования дигаплоидами S.tuberosum гибридов F1 между S.verrucosum и 1EBN дикими видами картофеля, из которой видно, что гибриды на основе Sv Sv-линий не уступают гибридам на основе S.verrucosum по способности скрещиваться в качестве материнских форм с дигаплоидами S.tuberosum. Предлагаемый способ получения гибридов между культурным картофелем Solanum tuberosum и 1EBN диплоидными дикими видами картофеля реализован следующим образом. Создание SvSv-линий В качестве исходного материала использовали 29 образцов самосовместимого дикого диплоидного вида Solanum verrucosum Schltdl. (2n=2x=24), семена которых были получены из Национального генного банка по картофелю США (NRSP-6). Эти образцы использовали в скрещиваниях в качестве опылителей с 13 фертильными вторичными дигаплоидами картофеля Solanum tuberosum L., 28 первичными дигаплоидами сортов Ласунак, Орбита, Альпинист (Беларусь). Atzimba. Ireri, Nortena. (Мексика), Ins (Польша), Granola (Германия), Agna (Нидерланды) из коллекции лаборатории генетики картофеля Института генетики и цитологии НАН Беларуси. Гибридизацию между дигаплоидами Solanum tuberosum и образцами Solanum verrucosum проводили следующим образом. Для проведения гибридизации растения высаживали в грунт оранжереи из культуры in vitro или путем посева семян в начале мая. Растения высаживали рядами: расстояние между рядами 70 см, между растениями в ряду 50 см. Режим выращивания растений: температура воздуха 20-25°C - днем, 15-20°C - ночью, влажность 70%, освещение естественное. Для стимуляции цветения растения, достигшие высоты 35-40 см, подвязывали к кольям, обнажали от почвы место отхождения корней от стебля. Образующиеся столоны обрывали или оставляли над поверхностью почвы. Для получения SvSv-линий растения дигаплоидов Solarium tuberosum использовали в качестве материнских форм. Накануне опыления проводили кастрацию бутонов, начинающих приобретать окраску (нижняя часть бутонов в это время имеет белый цвет, верхняя часть бутонов - цвет, характерный для цветков опыляемого образца), путем удаления пыльников с помощью пинцета. Также удаляли в соцветии все раскрытые цветки, полураскрытые и незрелые (зеленые или начинающие приобретать беловатую окраску) бутоны. Образцы Solarium verrncosum использовали в качестве опылителей. Сбор пыльцы проводили следующим образом: распустившиеся цветы собирали, подсушивали 1 сутки при комнатной температуре, затем вытряхивали из них пыльцу с помощью электрического вибратора, например, электробритвы типа Микма 100, 101, Эра и др. Собранную пыльцу переносили в лунки пластмассовых планшетов, применяемых для иммуноферментного анализа или пластиковые одноразовые чашки Петри диаметром 5 см. Опыление кастрированных цветков дигаплоидов Solanum tuberosum осуществляли путем погружения рыльца пестика в пыльцу. Использовали свежесобранную пыльцу или пыльцу, хранившуюся в морозильной камере при температуре - 10°C не более 5 дней. На опыленное соцветие вешали этикетку, в которой была указана дата опыления и наименование родительских форм. Применяли опыление смесью пыльцы или опыление пыльцой отдельных генотипов Solanum verrucosum (при опылении смесью пыльцы завязываемость семян значительно выше). Смесь пыльцы готовили непосредственно перед гибридизацией путем объединения и тщательного перемешивания в лунке равных объемов пыльцы разных образцов Solanum verrucosum. Пыльцевые трубки самосовместимого Solanum verrucosum не способны расти в пестиках самонесовместимых видов, в том числе дигаплоидов Solanum tuberosum (односторонняя несовместимость). Тем не менее, обнаружены отдельные, так называемые акцепторные, генотипы дигаплоидов Solanum tuberosum, которые могут завязывать семена при опылении Solanum verrucosum. Показано, что акцепторность Solanum tuberosum находится под контролем двух генов: гена акцепторности А и ингибитора акцепторности I (акцепторными являются генотипы A*ii) [7] (Фиг.1). В результате проведенной гибридизации между дигаплоидами Solarium tuberosum и Solanum verrucosum в изученной коллекции были обнаружены 4 акцепторных генотипа: 1) IGC 98/109-5: первичный дигаплоид сорта Nortena (Мексика); 2} гибрид Solanum tuberosum 881-28: [(Yul.2 × IGC-203/5.7) × Yul.2] × Yul.2; 3) гибрид 895-40: [(Yul.2 × IGC-203/5.7) × Yul.2] × Yul.2; 4) гибрид 885-21: [(Yul.2 × 203/5.7-20) × Yul.2] × А15, где Yu 1.2 - клон от свободного опыления первичного дигаплоида сорта Юбель, IGC-203/5.7 - вторичный дигаплоид на основе сорта Полесский розовый), А15 - клон S.chacoense Через месяц после опыления на завязавшиеся ягоды одевали марлевые мешочки и подвязывали их к стеблю материнского растения. Гибридные семена извлекали из созревших ягод с помощью скальпеля или пинцета, подсушивали и хранили в бумажных пакетах при комнатной температуре. Для посева их замачивали на сутки в 0,1% растворе гиббереллина и высевали в начале апреля в плошки с торфяной почвой «Двина», смоченной водой. Глубина заделки семян в почву 0,5 см. Растения гибридов F1 между акцепторными дигаплоидами S.tuberosum и S.verrucosum самоопыляли или проводили сестринские скрещивания (между разными растениями одной и той же гибридной комбинации) для получения гибридов F2. Целью получения поколения F2 является отбор гибридов, имеющих аллель Sv в гомозиготном состоянии (SvSv). Для большинства гибридов F1 была характерна высокая функциональная фертильность пыльцы, что обеспечило высокую результативность гибридизации с целью получения F2. Для дигаплоидов S.tuberosum характерна гаметофитная самонесовместимость. Тем не менее, среди них можно отобрать отдельные мутантные формы, способные к самоопылению. В настоящей работе в качестве материнских форм для получения гибридов с S.verrucosum использовано потомство самосовместимых дигаплоидов S.tuberosum, которые были отобраны нами ранее. Неслучайно поэтому отдельные гибриды F1 S.tuberosum × S.verrucosum оказались способными с высокой эффективностью завязывать семена при самоопылении. Те гибриды F1, которые были самонесовместимы, несложно было вовлечь в гибридизацию с сестринским формами: достаточно различие по одному из S-аллелей родительских форм, чтобы гибридизация была успешной. Семена, полученные в результате самоопыления гибридов F1, собирали и хранили как описано выше. Их высевали на следующий год для получения растений F2, среди которых проводили отбор Sv-гомозигот (S vSv-линий). Для выделения S v-гомозигот проводили отбор по данным анализа роста в пестиках гибридов пыльцевых трубок мексиканских видов S.bulbocastanum и S.pinnatisectum, несовместимых с S.tuberosum. Наблюдение за ростом пыльцевых трубок в пестиках осуществляли по методике, описанной в руководстве Абрамовой с соавторами [8] с помощью люминесцентного микроскопа «Люмам И-3». Пестики опыленных цветков (3-5 штук от каждой комбинации скрещивания) собирали через 48 часов после опыления и фиксировали в ацетоалкоголе (3:1). При приготовлении препаратов растительный материал дважды отмывался от фиксатора в дистиллированной воде, затем выдерживался в течение 20 минут в 10% спиртовом растворе КОН и окрашивался далее в растворе красителя анилинового синего (0,1% раствор анилинового синего в 0,05М K2HPO4, pH12), после чего приготавливались давленые препараты в 50% водном растворе глицерина. Рост пыльцевых трубок оценивали в баллах: 0 - ингибирование роста пыльцевых трубок непосредственно под рыльцем пестика, 1 - ингибирование роста пыльцевых трубок в верхней трети пестика, 2 - ингибирование на уровне середины - нижней трети пестика, 3 - завязи достигает 1-5 пыльцевых трубок, 4 - завязи достигает 15-30 пыльцевых трубок, 5 - завязи достигает толстый жгут пыльцевых трубок. В качестве SvSv-линий рассматривали растения, которые имели показатель роста пыльцевых трубок мексиканских видов равный пяти баллам. Подобную картину можно наблюдать при опылении такой пыльцой S.verrucosum, для которого характерно отсутствие пестичных S-РНКаз. Отобранные SvSv-линии непосредственно включали в гибридизацию с 1EBN дикими видами картофеля, либо получали клубни, из которых на следующий год выращивали растения SvSv-лнний для гибридизации с 1EBN дикими видами картофеля или дигаплоидами S.tuberosum (для дальнейшего беккроссирования). Получение промежуточных гибридов между посредником и 1EBN дикими видами картофеля Для вовлечения в гибридизацию с целью переноса в селекционный материал ценных генов от 1EBN диких видов картофеля проводили их гибридизацию разными образцами S.verrucosum (прототип) и отобранными SvSv-линиями (заявляемое решение). В качестве материала использовали 12 образцов S.verrucosum и 13 Sv Sv-линий (F2 S.tuberosum × S.verritcosum) (материнские формы), которые скрещивали с мужски фертильными образцами диплоидных 1EBN видов картофеля S.pinnatisectum, S.polyadenium, S.bulbocastanum. Семена образцов диких видов были получены из Национального генного банка по картофелю США (NRSP-6): S.verrncosum: PI 190170, PI 195171. PI 195172, PI 275256. PI 275257, PI 310966, PI 498010, PI 570643. PI 570644, PI 570645. PI 545745, PI 498060; S.pinnatisectum: PI 275231-275236, PI 537023; S.polyadenium: PI 230480. PI 275238. PI 347770. PI 498036; S.bulbocastanum: PI 243505, PI 243506. PI 243512. PI 243513. PI 255516. PI 275191. PI 275192. PH 98011. Выращивание растений и гибридизацию проводили, как описано выше, в 2009 г. В 2010 г семена полученных гибридов высевали, как описано выше, при этом учитывали всхожесть семян (количество взошедших семян/количество высеянных семян × 100%). У гибридных растений определяли показатели фертильности: способность к цветению, образование пыльцы и функциональную фертильность пыльцы. Для определения функциональной фертильности пыльцы (далее ФФП) проводили проращивание пыльцы in vitro по методике «висячей капли», предложенной N.Pallais [9]. Для проращивания пыльцы готовили среду следующего состава: сахароза - 200 г/л H3BO3 - 100 мг/л Ca(NO3)2×4H2 O - 300 мг/л MgSO4×7H2 O - 200 мг/л KNO3 - 100 мг/л TWEEN pH 5,5 По 3-5 капель среды (5 мкл каждая) наносились на поверхность влажных камер для проращивания. Пыльца равномерно распределялась по поверхности капель препаровальной иглой и выдерживалась 2-3 часа при комнатной температуре. После чего производился подсчет проросших пыльцевых зерен. Учитывали по 300 пыльцевых зерен с образца в нескольких полях зрения микроскопа (увеличение 600×. Функциональную фертильность пыльцы рассчитывали как: ФФП=частота проросших пыльцевых зерен/частота окрашенных пыльцевых зерен × 100%. В группу стерильных и низкофертильных гибридов относили те, которые не цвели, не образовывали пыльцу или имели ФФП ниже 5%. В группу среднефертильных относили генотипы, имеющие ФФП от 5 до 10%, а к высокофертильным относили генотипы с ФФП более 10%. Скрещивания промежуточных гибридов с дигаплоидами S.tuberosum проводили, как описано выше. Опыление гибридов проводили смесью пыльцы фертильных дигаплоидов S.tuberosum: 1) 881-10 - клон Библиографические данные: 1. Hermsen J.G.Th. Introgression of genes from wild species, including molecular and cellular approaches. / In: Potato Genetics (J.E.Bradshaw, G.R.Mackay eds.) - 1994. - CAB International. University Press, Cambridge. England. - P.515-539. 2. Jansky, S.H., Hamernik, A.J. 2009. The Use of Solanum Verrucosum as a Bridge Species. Genetic Resources and Crop Evolution. 56(8): 1107-1116. 3. Dinu I.I., Hayes R.J., Kynast R.G., Phillips R.L., Thill C.A. Novel inter-series hybrids in Solanum, section Petota // Theor Appl Genet. - 2005. - Vol.110. - P.403-415. Dinu I.I., Hayes R.J., Kynast R.G., Phillips R.L., Thill C.A. Novel inter-series hybrids in Solanum, section Petota // Theor Appl Genet. - 2005. - Vol.110. - P.403-415. 4. Маханько, О.В. Межвидовая несовместимость в диплоидной селекции картофеля // Земляробства i ахова раслiн. - 2008. - 5. Hanneman R.E. Jr. The reproductive biology of the potato and its implication for breeding // Potato Research. - 1999. - Vol.42/extra edition. - P.283-312. 6. Abdalla M.M., Hermsen J.G.Th. Plasmons and male sterility types in Solanum verrucosum and its interspecific hybrid derivatives // Euphytica. 1972. V.21. P.209-220. 7. Hermsen J.G.Th., Olsder J., Hoving E., Jansen P. Acceptance of self-compatible pollen from Solanum verrucosum in dihaploids from Solanum tuberosiim II Fertilization in higher plants. - 1974. - P.37-40. 8. Методические указания по цитологической и цитоэмбриологической технике (для исследования культурных растений) / Л.И. Абрамова, И.Н. Орлова, М.А. Вишнякова, Л.Н. Константинова, Л.И. Орел, В.Ф. Огородникова // Ленинград: ВИР, - 1981. - 118 с. 9. Pallais N., Fong N., Berrios D. Research on the physiology of potato sexual seed production // Innovative methods for propagating potatoes. CIP Rep. 28th Planning Conf: Proc. int. conf. - Lima: CIP, 1984. - P.149-168. Формула изобретенияСпособ получения гибридов между культурным картофелем Solanum tuberosum и 1EBN диплоидными дикими видами картофеля, включающий гибридизацию между lEBN диплоидными дикими видами картофеля и видом (линией)-посредником, не образующим пестичных РНК-аз, и последующее беккроссирование полученных гибридов дигаплоидами Solanum tuberosum, отличающийся тем, что получают и используют в качестве материнских форм при гибридизации между 1EBN диплоидными дикими видами картофеля и видом (линией)-посредником растения, отобранные в поколении F2 гибридов между дигаплоидами Solanum tuberosum и диким диплоидным видом картофеля Solanum verrucosum, несущие в гомозиготном состоянии S гены, привнесенные от Solanum verrucosum. Популярные патенты: 2062564 Способ оценки устойчивости растений к засухе северного и южного типа на ранних этапах онтогенеза ... Контроль растения, выращенные на воде при температуре 27В°С для засухи северного типа в течение 10 и 24 часов, для засухи южного типа 24 и 24 часа. Сроки экспозиции в контроле подбираются для получения одновозрастных растений. Преимуществом предлагаемого метода является его простота, возможность определения устойчивости к дефициту влаги в условиях максимально приближенных к естественным. Существенные отличия предлагаемого способа от аналогов (1,2,3,4) заключаются в том, что условия водного дефицита создаются растворами осмотиков (ПЭГ, сорбит) и определение ведется по показателю (изменение размеров корня), который можно определить с большой точностью с помощью инструментальных ... 2112341 Лапа плоскорежущая ... 3. Задняя часть 8 пластины 4 расположена эквидистантно боковой поверхности щеки 3. Верхний обрез 9 зигзагообразной пластины 4 смещен вперед по высоте относительно нижнего обреза 10 пластины 4. Нижний обрез 10 пластины 4 расположен параллельно задней кромке 11 односторонней лапы 2 и над ней. Длина l наклонной части зигзагообразной пластины 4 и длина L односторонней лапы 2 находятся в соотношении 1 : (8 - 10). Лапа плоскорежущая выполнена из листовой стали, развертка которой изображена на фиг. 6. Лапа плоскорежущая работает следующим образом. При обработке междурядий посевов пропашных культур на ровном поле стойку 1 располагают таким образом, чтобы расстояние от культурных растений ... 2182420 Устройство для перерезания стволов деревьев ... машиной подают к дереву, ствол 8 которого подлежит перерезанию до контакта ствола 8 с упорами 9 и гребенками 10, после чего при помощи силовых цилиндров 2, 3 поворачивают рычаги 6, 7 и зажимают ствол 8, прижимая его к упорам и гребенкам. Далее, используя привод с одним силовым цилиндром (фиг. 2), выдвигают его шток 20, который воздействует на заднюю поперечную сторону 15 полотна, перемещая последнее в направлении ствола 8 перерезаемого дерева. При этом кромки 16 взаимодействуют со стволом 8 и перерезают его в конце хода силового цилиндра (фиг.2). Перерезание может осуществляться продольными режущими лезвиями 17 кромок или режущими венцами 18 из корончатых резцов 19. В последнем ... 2262220 Способ возделывания кормовых культур в условиях астраханской области (варианты) ... из набора подсолнечника, мальвы, сорго и сои при зеленой спелости бобов сои, конца цветения корзинок подсолнечника и молочной спелости семян сорго. Сведения, подтверждающие возможность реализации заявленного способа, заключаются в следующем.В условиях Нижней Волги, при большом дефиците почвенной влаги и частых атмосферных и почвенных засухах возделывание сои возможно только в условиях орошения на любых типах почв.Технология возделывания сои, как основной культуры в смесях, в условиях аридного земледелия поэтапно отрабатывалась нами на протяжении 10 лет. Вначале отрабатывались элементы агротехники возделывания и режимы поливов, затем велось изучение, подбор новых сортов, созданных ... 2142331 Устройство для гомогенизации и гомогенизирующая головка ... в следующей последовательности: стержень 20 с герметизирующим кольцом 26 продевается в отверстие стенки 28 таким образом, чтобы направляющая планка 21 наделась на направляющие стержни 25. Стенка 28 закрепляется винтами на торце корпуса 14. Далее по очереди накручиваются опорная гайка 24 на стенку 28, и гайка 22 на стержень 20. Гайка 22 накручивается до тех пор, пока проточная канавка на ней не совпадает с поверхностью опорной гайки 24. После этого в проточную канавку гайки 22 вставляется фиксирующая шайба 23 и прикрепляется винтами к опорной гайке 24. Затем с другого торца корпуса 14 вставляется щелевое сопло с патрубком 8 так, чтобы продольная ось его щели совпала с заостренной ... |
Еще из этого раздела: 2201069 Травяное покрытие на основе гибкого полотна 2132610 Устройство обогрева сельскохозяйственных животных и птицы 2059362 Установка для выращивания мидий 2048744 Устройство для регулирования температуры воздуха в теплице 2038763 Регулятор вакуума 2471341 Стойло, устройство в стойле и способ монтажа указанного устройства 2121263 Способ лесоводственной оценки технологического комплекса машин 2236122 Устройство для содержания животных 2232490 Машина для обработки почвы 2414114 Зерноуборочный комбайн |
Изобретения в сельском хозяйстве
Обработка почвы в сельском и лесном хозяйствах
Посадка, посев, удобрение
Уборка урожая, жатва
Обработка и хранение продуктов полеводства и садоводства
Садоводство, разведение овощей, цветов, риса, фруктов, винограда, лесное хозяйство
Новые виды растений или способы их выращивания
Производство молочных продуктов
Животноводство, разведение и содержание птицы, рыбы, насекомых, рыбоводство, рыболовство
Поимка, отлов или отпугивание животных
Консервирование туш животных, или растений или их частей
Биоцидная, репеллентная, аттрактантная или регулирующая рост растений активность химических соединений или препаратов
Хлебопекарные печи, машины и прочее оборудование для хлебопечения
Машины или оборудование для приготовления или обработки теста
Обработка муки или теста для выпечки, способы выпечки, мучные изделия
|
|
||
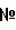 10 комбинации скрещивания [Yu 1.2 × (Yul.2 × IGC-203/5.7-20)]-5 × А15; 2) 882-5 - клон
10 комбинации скрещивания [Yu 1.2 × (Yul.2 × IGC-203/5.7-20)]-5 × А15; 2) 882-5 - клон