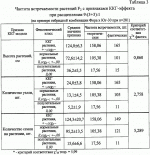
Способ получения высокопродуктивных растений- самоопылителей на основе эффекта закрепленного гетерозисаПатент на изобретение №: 2254709 Автор: Зеленцов С.В. (RU), Кочегура А.В. (RU), Петибская В.С. (RU), Мошненко Е.В. (RU) Патентообладатель: Государственное научное учреждение Всероссийский научно-исследовательский институт масличных культур им. В.С. Пустовойта Российской сельскохозяйственной академии (RU) Дата публикации: 27 Июня, 2005 Начало действия патента: 11 Декабря, 2003 Адрес для переписки: 350038, г.Краснодар, ул. Филатова, 17, ГНУ ВНИИМК РАСХН, патентная группа Изображения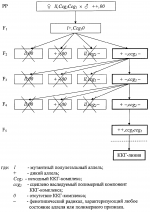 Изобретение относится к области сельского хозяйства, в частности селекции растений. Подбирают родительские формы по гетерозису, на основе гибридологического анализа по наличию гомозиготных полулетальных мутаций в F2 у одной из родительских форм. Далее проводят скрещивание, отбирают гибриды по гетерозису, оценивают и размножают гомозиготные формы с максимальным ККГ-эффектом. До проведения отбора формируют морфологическую ККГ-модель путем проведения подробного морфологического описания растений F 1, полученных от скрещивания подобранных родителей, для выявления ККГ-эффекта по данной полулетальной мутации, а отбор гибридов по гетерозису осуществляют путем выделения в F2 растения с признаками ККГ-модели и высева семян каждого отобранного растения на индивидуальной делянке. В F3 и F4 отбор проводят аналогичным способом, как в F 2, а в F5, выделяют гомозиготные по селектируемым признакам формы с максимальным ККГ-эффектом. Способ позволяет получить высокожизнеспособные формы растений-самоопылителей и тем самым повысить продуктивность создаваемых сортов у самоопыляющихся культур. 1 ил., 3 табл. Изобретение относится к сельскому хозяйству и может быть использовано в селекции растений. Известно, что гетерозис - свойство гибридов превосходить по определенным признакам лучшую из родительских форм. Максимально проявляется оно в первом поколении гибридов, а в последующих - постепенно затухает. Закрепить его в потомстве ученым-селекционерам долго не удавалось. Впервые метод закрепления гетерозиса в потомстве был разработан на тутовом шелкопряде. Генетические же закономерности возникновения и затухания гетерозиса в потомстве растений такие же, как и у животных. Поэтому известные способы закрепления гетерозиса на животных объектах (на тутовом шелкопряде) могут быть использованы в качестве аналогов для растительных объектов. Известен способ селекции тутового шелкопряда, включающий использование в качестве родителей форм с полулетальными мутациями, длительного инбридинга (инцухта), отбора из инбредных линий высоко жизнеспособных форм, несущих полулетальный ген l, который нейтрализован комплексом скоординировано действующих компенсационных генов (ККГ), вовлечения таких форм в гибридизацию, оценки их общей (ОКС) и специфической (СКС) комбинационной способности и отбора инбредных форм с высокой ОКС и СКС, обеспечивающих мощный гетерозис за счет положительного аддитивного действия ККГ на жизнеспособность организмов типа l/+ на фоне гетерозиготного подавления негативного эффекта полулетальных генов (Струнников В.А. Новая гипотеза гетерозиса и ее научное и практическое значение. Вестник с.-х. науки, 1983, №1(316), С.34-40). Недостатком известного способа является то, что увеличение продуктивности организмов за счет мощного проявления гетерозиса обеспечивается только в первом гибридном поколении F1. В последующих поколениях наблюдается уменьшение эффекта гетерозиса за счет перехода полулетальных генов в гомозиготное (l) состояние, а также нарушения координации ККГ вплоть до его полного распада за счет последующего внутрипопуляционного неконтролируемого перекрестного скрещивания. При выведении линий с высокой комбинационной способностью на основе ККГ требуется длительный период времени, обусловленный необходимостью проведения нескольких беккроссных скрещиваний для выведения из генома полулетальных генов и последующего перевода беккроссной линии в гомозиготное состояние по нормальному (+) аллелю стандартными способами. В целом, использование известного способа селекции с длительными периодами отбора высокожизнеспособных инбредных линий и многократным беккроссированием пригодно только для организмов, имеющих короткие циклы онтогенетического развития и дающих несколько поколений в год, и существенно ограничивает его применение в селекции растений, дающих одну генерацию в год. Промышленное использование гибридов F1, полученных известным способом, возможно только на перекрестие опыляющихся видах растений. Известен также способ закрепления основанного на ККГ-эффекта гетерозиса в последующих поколениях организмов. Для этого подбором родительских форм получают высокогетерозисные гибриды, от которых получают серию абсолютно гомозиготных потомков методом моноспермического андрогенеза. У растений абсолютные гомозиготы получают путем стимулирования эмбрионального развития гаплоидной пыльцы с последующим превращением ее зародышевых клеток в диплоидные, развивающиеся в жизнеспособные фертильные растения. После 5-6 беккроссов абсолютно гомозиготных особей с исходным гибридом очищенный от полулетальных генов гибрид массово размножают путем внутривидового скрещивания. При этом достигается почти полное удаление леталей и полулеталей из генотипа, в то же время сохраняется часть генов, обеспечивающих гетерозис в исходном гибриде (Струнникова Л.В. Струнников В.А. Способ закрепления гетерозиса гибридов в последующих поколениях. Патент РФ №2153253, А 01 К 67/00, 67/04). Недостатком известного способа, взятого в качестве прототипа, является то, что для сохранения гетерозиса в ряду последовательных поколений необходимо проведение 5-6 беккроссов полученных гибридов с их абсолютно гомозиготными потомками (сыновьями). Для выведения абсолютно гомозиготных потомков требуется наличие метода их получения, основанного на моноспермическом андрогенезе. При этом у тутового шелкопряда неосемененных самок любой породы облучают Задача, решаемая изобретением, состоит в повышении продуктивности растений-самоопылителей за счет использования суммарного эффекта комплекса компенсирующих генов (ККГ-комплекса) на фоне отсутствия ККГ-образующей полулетальной мутации в гомозиготном потомстве. Цель изобретения - передача от родительской формы растения-самоопылителя, несущего полулетальную мутацию, накопленного в филогенезе блока коадаптированных генов ККГ-комплекса и закрепление его в гомозиготном потомстве. Технический эффект достигается тем, что в известном способе закрепления гетерозиса гибридов в последующих поколениях, включающем подбор родительских форм по гетерозису, определяемому большим количеством скоординированных в своем действии благоприятных генов (ККГ-комплексом), их скрещивание, отбор гибридов по гетерозису, оценку и размножение гомозиготных форм с максимальным ККГ-эффектом, согласно изобретению подбор родительских форм осуществляют на основе гибридологического анализа по наличию гомозиготных полулетальных мутаций в F2 у одной из родительских форм, формируют морфологическую ККГ-модель путем подробного морфологического описания растений F1, полученных от скрещивания подобранных родителей, для выявления ККГ-эффекта по данной полулетальной мутации, а отбор гибридов по гетерозису осуществляют следующим образом: в F2 выделяют растения с признаками морфологической ККГ-модели и высевают семена каждого отобранного растения на индивидуальной делянке, в F3 и F4 поступают также, как и в F2, а в F5 выделяют гомозиготные по селектируемым признакам формы с максимальным ККГ-эффектом. Сопоставительный анализ заявляемого технического решения с прототипом позволяет сделать вывод, что заявляемый способ отличается от известного наличием новой операции (формирование морфологической ККГ-модели путем подробного морфологического описания растений F1, полученных от скрещивания подобранных родителей, для выявления ККГ-эффекта по данной полулетальной мутации), а также порядком выполнения известной операции “подбор родительских форм” (эту операцию осуществляют не подбором родителей, обеспечивающих наиболее высокогетерозисные гибриды, как это изложено в прототипе, а на основе гибридологического анализа по наличию гомозиготных полулеталей в F2) у одной из родительских форм, и порядком выполнения известной операции “отбор гибридов по гетерозису”: в прототипе эту операцию осуществляют путем сравнительных испытаний серии индивидуальных гибридов и выбором 10 лучших по гетерозису индивидуальных гибридов, а в заявляемом способе по другому: в F2 выделяют растения с признаками морфологической ККГ-модели и высевают семена каждого отобранного растения на индивидуальной делянке, в F2 и F3 поступают так же, как и в F2, а в F5 выделяют гомозиготные по селектируемым признакам формы с максимальным ККГ-эффектом. Таким образом, заявляемый способ соответствует критерию патентоспособности “НОВИЗНА”. Исследуя уровень техники в процессе проведения патентного поиска по всем видам сведений, общедоступных в печати, обнаружили, что способ получения высокопродуктивных растений-самоопылителей с использованием эффекта закрепленного гетерозиса, включающий заявленную совокупность признаков, для специалиста в области селекции явным образом не следует из известного на сегодня существующего уровня техники. Исследованиями на протяжении ряда лет установлено, что геномы живых организмов, в том числе геномы растений-самоопылителей в филогенезе, подвергаются постоянному воздействию как естественных, так и антропогенных мутагенных факторов и накапливают мутации. Подавляющее большинство мутаций оказывает негативный эффект в онтогенезе. Полулетальные и субвитальные мутации, не затрагивающие существенно репродуктивные функции организма, за счет горизонтального переноса генов и рекомбинационных процессов в мейозе способствуют постепенному накоплению доминантных и полудоминантных генов, компенсирующих негативный эффект мутаций. Компенсационный эффект сложившихся блоков коадаптированных генов ККГ-комплекса пропорционален вредоносности мутации; как правило, направлен на улучшение экспрессии признаков, подавляемых мутацией, и наиболее выражен у организмов - носителей полулетальных мутаций. В ряде случаев ККГ-комплекс полностью нейтрализует негативный эффект полулетальной мутации, при этом генотипы, несущие нейтрализованную ККГ-комплексом мутацию, могут быть визуально и морфологически неотличимы от здоровых организмов. При расщеплении гибридов, образовавшихся с участием родительских генотипов - источников ККГ-комплексов, образуются генотипические классы с отсутствием полулетальной мутации и наличием активных полимерных компонентов исходного ККГ-комплекса, обеспечивающим сохраняющийся в ряду поколений гетерозис (ККГ-эффект). В таких генотипах при отсутствии вредной мутации компенсирующий эффект коадаптированного блока генов ККГ-комплекса или его сцеплено наследуемых активных компонентов реализуется в положительном аддитивном воздействии на весь организм, повышая его общую жизнеспособность и улучшая его морфологические показатели. Гомозиготные гибридные генотипы - носители ККГ-комплекса (ККГ-линии), отличающиеся улучшенными хозяйственно-ценными признаками, могут использоваться как в качестве исходного материала при селекции растений, так и в качестве полноценных сортов. На основании вышеизложенного можно сделать вывод о соответствии заявляемого технического решения критерию патентоспособности “ИЗОБРЕТАТЕЛЬСКИЙ УРОВЕНЬ”. Заявляемое техническое решение соответствует и критерию патентоспособности “ПРОМЫШЛЕННАЯ ПРИМЕНИМОСТЬ”, так как оно может быть использовано в сельском хозяйстве, и, кроме того, в описании изобретения представлены средства и методы, с помощью которых возможно осуществление технического решения в том виде, как оно охарактеризовано в формуле изобретения. Способ осуществляют следующим образом. Начинают с подбора родительских форм по гетерозису, определяемому большим количеством скоординированных в своем действии благоприятных генов (ККГ-комплекса). Наличие скомпенсированной полулетали в геноме определяют гибридологическим методом по наличию в F2 гомозигот по полулетальной мутации у одной из родительских форм. При этом в качестве другого родителя берут любую генетически неродственную форму того же вида с диким (не мутантным) аллелем по месту полулетальной мутации первого родителя и отсутствием экспрессии ККГ-эффекта. Проводят скрещивание подобранных родительских форм любым известным способом. Формируют морфологическую ККГ-модель путем подробного морфологического описания растений F1, полученных от скрещивания подобранных родителей, для выявления ККГ-эффекта по данной полулетальной мутации. Отбор гибридов по гетерозису осуществляют следующим образом: в F2 выделяют растения с признаками морфологической ККГ-модели и высевают семена каждого отобранного растения на индивидуальной делянке. В F3 и F4 поступают так же, как и в F2. А в F5 выделяют гомозиготные по селектируемым признакам формы с максимальным ККГ-эффектом. Выделенные гомозиготные формы с максимальным ККГ-эффектом оценивают по стандартным методикам и размножают. Пример осуществления способа. Были проанализированы расщепляющиеся популяции гибридных комбинаций сои за предыдущие 10 лет (всего около 700 комбинаций) и выделены родительские формы, у которых в F2 выщеплялись полулетальные растения. Таким образом, в качестве предполагаемых источников ККГ-комплексов были выделены сортообразцы сои Фора и Т-245 типа ll,Ccg1Ccg1, (где l - полулетальная мутация; Ccg1 - исходный комплекс компенсационных генов), которые были взяты в качестве исходных форм для последующего гибридологического анализа. Выделенные сортообразцы сои Фора и Т-245 были вовлечены в скрещивания с набором из 3-х сортов, не имеющих в своих геномах полулетальных мутаций (табл.1). За нулевую гипотезу принимали дигенное расщепление с полным доминированием и сцепленным наследованием активных компонентов ККГ-комплекса с соотношением фенотипических классов 9:3:3:1. При этом частота встречаемости рецессивных полулеталей теоретически должна составлять 1/16, а частота встречаемости растений без видимых признаков полулетальности - 15/16. Анализ типов феноклассов в расщепляющейся популяции F 2 выявил наличие полулетальных растений при скрещивании сортов Фора и Т-245 со всеми сортами. При скрещивании контрольного сорта Астра были обнаружены только единичные растения полулетального морфотипа. Гибридологический анализ частот фенотипических классов показал, что во всех гибридных комбинациях с участием сортов Фора и Т-245 обнаруживается близкое к 15:1 соотношение фенотипически нормальных и полулетальных растений. Более низкие, по сравнению с теоретическими, фактические значения Таким образом, в геноме сортов сои Фора и Т-245 было подтверждено наличие полулетальных мутаций, а отсутствие у них видимых признаков снижения жизнеспособности или угнетения свидетельствовало о наличии активного ККГ-комплекса, эффективно нейтрализующего полулетальную мутацию. Сорта сои Фора и Т-245 в качестве источников ККГ-комплексов были скрещены с неродственными сортами сои того же вида (с генотипами типа ++,00, где + - дикий (не мутантный) аллель, а 0 - отсутствие ККГ-комплекса), адаптированными к условиям произрастания в зоне осуществления селекции. В поколении F1 исходная рецессивная гомозиготная мутация ll перешла в гетерозиготное состояние l+ и фенотипически не проявлялась, а одинарный, но скоординированный блок генов ККГ-комплекса оказал аддитивное положительное влияние на весь гибридный организм, обеспечивая повышенную жизнеспособность (мощный гетерозис). Исследовали и описали фенотип растений F1 типа l+,Ccg1 0 с одинарной дозой ККГ-комплекса, который в дальнейшем послужил морфологической моделью для оценки экспрессии активных компонентов коадаптированного блока генов данного ККГ-комплекса при отборе форм типа ++,ccg2ccg2 (где ccg2 - сцеплено наследуемые компоненты ККГ-комплекса) (Фиг.1). Полученные семена F1 высевали отдельными делянками по комбинациям в оптимальные для культуры сроки. Рядом на отдельных рядках выращивали растения обоих родительских сортов. В течение вегетации проводили фенологические наблюдения. В фазу физиологической спелости семян гибридных растений F1 проводили биометрические измерения, определяя высоту растений, количество узлов, бобов, семян, массу 1000 семян. Эти же параметры определяли у родительских сортов (табл. 2). Как следует из анализа данных табл. 2, экспрессия одинарной дозы ККГ-комплекса растений F1 на фоне подавленной в гетерозиготе полулетали заключалась в трансгрессивном увеличении высоты растений, количества узлов на главном стебле и большем количестве семян. Эти параметры составили основу морфологической ККГ-модели по данной гибридной комбинации. Все полученные на растениях F1 семена F 2 высевали на следующий год отдельными делянками по комбинациям. В фазу физиологической спелости растений в расщепляющихся комбинациях F2 проводили морфологическую оценку по основным параметрам сформированной в пределах гибридной комбинации морфологической ККГ-модели. В расщепляющейся гибридной популяции F 2 выделяли следующие фенотипические классы: - гомозиготы по полулетали типа 11,00 полулетального фенотипа; - трансгрессивные (гетерозисные) гибридные формы фенотипического класса + -ccg 2-, близкие по морфотипу к эталонным растениям F1 , у которых присутствовали активные компоненты ККГ-комплекса и отсутствовала ККГ-образующая полулеталь; - гибридные формы - носители как полулетали, так и элементов ККГ-комплекса фенокласса ll,ccg2-, а также гибридные формы с отсутствием ККГ-комплекса фенокласса + -00, норма реакции которых не выходила за крайние значения соответствующих признаков обеих родительских форм. В гибридной популяции F2 дигенное расщепление по признакам полулетальности и ККГ-эффекта должно образовывать фенотипические классы: + -,ccg2-; ll,ccg2 -; +-,00 и ll,00 в соотношении 9:3:3:1. Однако из-за морфологической схожести и непрерывной изменчивости морфологических признаков с нормой реакции в пределах крайних значений соответствующих признаков обеих родительских форм феноклассы ll,ccg2-, и + -,00 были объединены в одну группу с теоретической частотой (3/16+3/1 б)=37,5%. Таким образом, в расщепляющейся гибридной популяции F2 выделяли, подсчитывали и оценивали по модельным признакам растения следующих 3-х фенотипических классов: гомозиготы по полулетали полулетального фенотипа; трансгрессивные гибридные формы фенокласса, близкие по морфотипу к ККГ-модели; гибридные формы феноклассов ll,ccg2-, и +-,00 (табл. 3). Как следует из табл. 3, выделение в F1 морфологических признаков для описания ККГ-модели по данной комбинации оказалось эффективным при отборе ККГ-форм в F2. Это позволило разделить все растения F2 в этой комбинации на 3 морфологически различающиеся группы. Все полулетальные растения типа ll,00, а также гибридные формы фенотипических классов ll,ccg2 -, и +-,00 выбраковывали. Растения фенокласса +-,ccg2 -, с положительными трансгрессиями по признакам ККГ-модели, составившие в гибридной комбинации Фора × Юг-30 53,0-58,7% от общего числа анализируемых растений, были пересеяны для получения следующего F3 поколения. В гибридных поколениях F3 -F4 повторяли процедуру визуальной и морфологической оценки растений, отбирая трансгрессивные растения фенокласса +-,ccg2-, и выбраковывая гибридные формы феноклассов ll,00; ll,ccg2- и + -, 00. Потомство отобранных в F4 ККГ-растений высевали на индивидуальных делянках для их оценки на генетическую однородность. В F5 отбирали нерасщепляющиеся ККГ-линии типа ++,ccg2ccg 2 с положительными трансгрессиями по основным хозяйственно-ценным признакам. Отобранные ККГ-линии оценивали по продуктивности и другим хозяйственно-ценным признакам по стандартным методикам. Лучшие по продуктивности и другим хозяйственно-ценным признакам ККГ-линии размножали стандартными методами для промышленного использования. Данный способ применим для создания высокопродуктивных растений у любых других самоопыляющихся культур. Подбор родительских форм на основе гибридологического анализа по наличию гомозиготных полулетальных мутаций в F2 у одной из родительских форм обеспечивает подтверждение наличия в их геноме полулетальных мутаций, а отсутствие у исходных сортообразцов видимых признаков снижения жизнеспособности свидетельствует о наличии эффективно нейтрализующего полулетальные мутации ККГ-комплекса. Такие сортообразцы могут использоваться как источники ККГ в последующих скрещиваниях, обеспечивая, таким образом, передачу от родительской формы растения-самоопылителя, несущего полулетальную мутацию, накопленного в филогенезе блока коадаптированных генов ККГ-комплекса и, в конечном итоге, повышение продуктивности растений-самоопылителей за счет использования суммарного эффекта комплекса компенсирующих генов (ККГ-комплекса) на фоне отсутствия ККГ-образующей полулетальной мутации в гомозиготном потомстве. Формирование морфологической ККГ-модели путем подробного морфологического описания растений F1, полученных от скрещивания подобранных родителей для выявления ККГ-эффекта по данной полулетальной мутации, обеспечивает выявление экспрессии одинарной дозы ККГ-комплекса в F1 на фоне подавленной в гетерозиготе полулетали и определение параметров, составляющих основу морфологической ККГ-модели по данной гибридной комбинации. Отбор гибридов по гетерозису заявленным путем, т.е. когда в F2 выделяют растения с признаками морфологической ККГ-модели, обеспечивает разделение гибридной популяции на морфологически различающиеся феногруппы, обеспечивая, таким образом передачу от родительской формы растения-самоопылителя, несущего полулетальную мутацию, накопленного в филогенезе блока коадаптированных генов ККГ-комплекса. Высев семян каждого отобранного растения с признаками морфологической ККГ-модели на индивидуальной делянке осуществляют для получения следующего F3 поколения с признаками морфологической ККГ-модели. Проведение такого же отбора растений с ККГ-эффектом в F3 и F4 , как и в F2, обеспечивает последовательное освобождение гибридной популяции от носителей полулетальных мутаций и увеличение доли растений с выраженным ККГ-эффектом, таким образом обеспечивая закрепление накопленного в филогенезе блока коадаптированных генов ККГ-комплекса в гомозиготном потомстве и повышение продуктивности растений-самоопылителей. Выделение в F5 гомозиготных по селектируемым признакам форм с максимальным ККГ-эффектом обеспечивает получение исходных ККГ-линий для проведения дальнейших оценок и передачи их в качестве сортов на государственное сортоиспытание, обеспечивая закрепление накопленного в филогенезе блока коадаптированных генов ККГ-комплекса в гомозиготном потомстве. Таким образом, обеспечивается повышение продуктивности растений-самоопылителей за счет использования суммарного эффекта комплекса компенсирующих генов (ККГ-комплекса) на фоне отсутствия ККГ-образующей полулетальной мутации в гомозиготном потомстве. Таким образом, заявляемым изобретением обеспечивается достижение поставленной цели и решение поставленной задачи. Формула изобретенияСпособ получения высокопродуктивных растений-самоопылителей с использованием эффекта закрепленного гетерозиса (ККГ-эффекта), включающий подбор родительских форм по гетерозису, определяемому большим количеством скоординированных в своем действии благоприятных генов (ККГ-комплекса), их скрещивание, отбор гибридов по гетерозису, оценку и размножение гомозиготных форм с максимальным ККГ-эффектом, отличающийся тем, что подбор родительских форм осуществляют на основе гибридологического анализа по наличию гомозиготных полулетальных мутаций в F2 у одной из родительских форм, формируют морфологическую ККГ-модель путем подробного морфологического описания растений F1, полученных от скрещивания подобранных родителей, для выявления ККГ-эффекта по данной полулетальной мутации, а отбор гибридов по гетерозису осуществляют следующим образом: в F2 выделяют растения с признаками морфологической ККГ-модели и высевают семена каждого отобранного растения на индивидуальной делянке, в F3 и F4 поступают так же, как и в F2, а в F5 выделяют гомозиготные по селектируемым признакам формы с максимальным ККГ-эффектом. MM4A Досрочное прекращение действия патента из-за неуплаты в установленный срок пошлины за поддержание патента в силе Дата прекращения действия патента: 12.12.2008 Дата публикации: 10.05.2011 Популярные патенты: 2111642 Высевающий аппарат ... по аналогии с отверстиями в днище 25 высевающего устройства 4. Пластины 39 соединены между собой поперечными планками 41 и 42. Поперечная планка 42 через пружину 43 прижимается к упорам 44. Пружина 43 другим своим концом связана с кронштейном 45, закрепленным на днище 25 высевающего устройства 4. Поперечная планка 41 через соединительное звено 46 связывается с любым исполнительным механизмом (электромагнитное реле, гидроцилиндр или же механическая связь с рамой сошниковой секции), который на схеме не показан. В рабочем режиме высевающего устройства 4 поперечная планка 42 пружиной 43 фиксируется упорами 44 так, что продолговатые высевные отверстия 26 днища 25 открыты. При разворотах ... 2215407 Способ создания исходного материала для селекции растений ... в течение ряда поколений. Таким образом, заявляемый способ соответствует критерию патентоспособности "НОВИЗНА". Исследуя уровень техники в процессе проведения патентного поиска по всем видам сведений, общедоступных в печати, мы обнаружили, что способ создания исходного материала для селекции растений, включающий заявленную нами совокупность признаков, для специалиста в области селекции явным образом не следует из известного на сегодня существующего уровня техники. Нашими исследованиями на протяжении ряда лет выявлено, что в природе с определенной частотой постоянно осуществляется спонтанная полиплоидизация высших растений. Полиплоидизация приводит к образованию поливалентов ... 2218756 Способ изготовления антипаразитарного ошейника ... стадии с одинаковыми температурными режимами при соотношении сухих и жидких исходных компонентов на вспомогательной стадии,%: Сухие: Сэвилен - 74,00-75,00 Стеариновокислый кальций - 0,60-2,95 Органический краситель - 0,10 Жидкие: Диазинон, 60% к.э. (концентрированная эмульсия) - 22,50-23,85 Масло мяты перечной - 0,45 сухие и жидкие компоненты смешивают, обеспечивая смачивание каждой гранулы термопластичного полимера жидкими компонентами с захватом низкомолекулярных твердых частиц с последующим превращением на стадии вспомогательной экструзии в гранулят, на стадии основной экструзии полученный гранулят перерабатывают в ленту при температурных режимах в материальном цилиндре ... 2312500 Способ защиты смородины от вредителей и болезней ... и агрохимикатов, разрешенных к применению на территории Российской Федерации, 2005 г.). Небольшой ассортимент биопестицидов, применяемых на ягодных кустарниках, которые обладают ограниченной возможностью, малоэффективен при влажной и нетеплой погоде (ниже 18-20°С), и приемлемой эффективностью подавляет не всех вредителей, особенно плохо скрытно и полускрытно живущих, и может успешно применяться только в комплексе дополняющих их действие защитных мероприятий.При этом обработку проводят не только до цветения и после сбора урожая, но и после цветения и в период формирования - созревания урожая (что чревато опасностью отравления ягод остатками ядохимикатов), поскольку ряд ... 2500104 Способ приготовления препарата костной ткани и набор для его осуществления ... гистологические препараты как при применении рутинных методов окрашивания (окрашивание гематоксилин-эозином), так и при применении иммуноморфологических методов окрашивания.При приготовлении препарата костной ткани используют набор, содержащий молекулярный фиксатор FineFix на спиртовой основе, содержащий FineFix и 96° спирт в соотношении 1:2,5, концентрированный раствор декальцинацинатора и рабочие растворы для контроля полноты декальцинации: 25% раствор аммиака и насыщенный раствор оксалата аммония. Сущность предлагаемого способа поясняется фигурами - гистологическими препаратами костной ткани, полученными по предлагаемому способу. Так, на фиг.1 показан ... |
Еще из этого раздела: 2015633 Способ переработки отходов животноводческих комплексов и устройство для его осуществления 2027341 Бункер для сыпучих материалов 2025945 Способ выращивания насаждений сосны 2108700 Способ оценки горных сенокосов и пастбищ 2167510 Способ и устройство для изготовления круглых тюков соломы или подобного материала с пленочным защитным покрытием 2114555 Способ электродиагностики вымени крупного рогатого скота и устройство для его осуществления 2475025 Средство для обработки семян зерновых и зернобобовых культур, пораженных фузариозом 2092036 Способ микроразмножения стевии stevia rebaudiana l. 2403703 Способ интенсификации роста растений 2091023 Способ защиты растений от заболеваний, вызванных нематодами |
Изобретения в сельском хозяйстве
Обработка почвы в сельском и лесном хозяйствах
Посадка, посев, удобрение
Уборка урожая, жатва
Обработка и хранение продуктов полеводства и садоводства
Садоводство, разведение овощей, цветов, риса, фруктов, винограда, лесное хозяйство
Новые виды растений или способы их выращивания
Производство молочных продуктов
Животноводство, разведение и содержание птицы, рыбы, насекомых, рыбоводство, рыболовство
Поимка, отлов или отпугивание животных
Консервирование туш животных, или растений или их частей
Биоцидная, репеллентная, аттрактантная или регулирующая рост растений активность химических соединений или препаратов
Хлебопекарные печи, машины и прочее оборудование для хлебопечения
Машины или оборудование для приготовления или обработки теста
Обработка муки или теста для выпечки, способы выпечки, мучные изделия
|
|
||
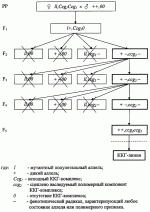
 -лучами в дозе 80 кр. Затем самки спариваются с самцами индивидуальных гибридов. Отложенные яйца в возрасте 60-80 минут после откладки при температуре 25°С прогревают 210 минут в воде, нагретой до 38°С. При этом абсолютные гомозиготы в подавляющем большинстве погибают на разных стадиях развития. Выживают только те гомозиготы, которым в ходе мейоза не досталось совсем или досталось очень мало вредных генов, в основном, слабого действия. У растений абсолютные гомозиготы получают путем стимулирования эмбрионального развития гаплоидной пыльцы с последующим превращением ее зародышевых клеток в диплоидные, развивающиеся в жизнеспособные фертильные растения. В целом применение известного способа, выбранного в качестве прототипа, существенно ограничивает выведение высокопродуктивных форм из-за обязательного включения в селекционный процесс андрогенетических потомков и длительного процесса беккроссирования. При этом успешный моноспермический андрогенез известен только у ограниченного числа видов растений и требует применения ряда последовательных биотехнологических операций с культурой пыльников, а многократные беккроссы с оценкой по потомству требуют длительного времени. Кроме этого, промышленное использование форм с закрепленным гетерозисом, полученных известным способом, возможно только на перекрестие опыляющихся видах растений.
-лучами в дозе 80 кр. Затем самки спариваются с самцами индивидуальных гибридов. Отложенные яйца в возрасте 60-80 минут после откладки при температуре 25°С прогревают 210 минут в воде, нагретой до 38°С. При этом абсолютные гомозиготы в подавляющем большинстве погибают на разных стадиях развития. Выживают только те гомозиготы, которым в ходе мейоза не досталось совсем или досталось очень мало вредных генов, в основном, слабого действия. У растений абсолютные гомозиготы получают путем стимулирования эмбрионального развития гаплоидной пыльцы с последующим превращением ее зародышевых клеток в диплоидные, развивающиеся в жизнеспособные фертильные растения. В целом применение известного способа, выбранного в качестве прототипа, существенно ограничивает выведение высокопродуктивных форм из-за обязательного включения в селекционный процесс андрогенетических потомков и длительного процесса беккроссирования. При этом успешный моноспермический андрогенез известен только у ограниченного числа видов растений и требует применения ряда последовательных биотехнологических операций с культурой пыльников, а многократные беккроссы с оценкой по потомству требуют длительного времени. Кроме этого, промышленное использование форм с закрепленным гетерозисом, полученных известным способом, возможно только на перекрестие опыляющихся видах растений. 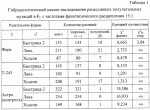
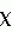 2 свидетельствуют о справедливости нулевой гипотезы при 0,05 уровне значимости.
2 свидетельствуют о справедливости нулевой гипотезы при 0,05 уровне значимости.