Способ получения неприродной трансгенной мыши с дефицитом гена рецептора 2 кортиколиберина и ее использованиеПатент на изобретение №: 2251838 Автор: ЛИ Куо-Фен (US), БЭЙЛ Трейси Л. (US), СМИТ Джордж В. (US), ВЭЙЛ Уайли (US) Патентообладатель: РИСЕРЧ ДИВЕЛОПМЕНТ ФАУНДЕЙШН (US) Дата публикации: 10 Марта, 2004 Начало действия патента: 14 Июля, 2000 Адрес для переписки: 129010, Москва, ул. Б.Спасская, 25, стр.3, ООО "Юридическая фирма Городисский и Партнеры", пат.пов. Е.Е.Назиной Изображения  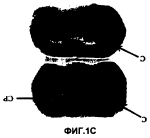 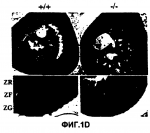 Изобретение относится к области генетической инженерии, в частности к способу получения неприродной трансгенной мыши с дефицитом гена рецептора 2 кортиколиберина (CRFR2). Способ предусматривает выделение геномного клона, содержащего локус гена CRFR2. Далее проводят конструирование направляющего вектора, в котором экзоны 10, 11 и 12 гена CRFR2 заменены кассетой селективного маркерного гена, и его перенос в эмбриональные стволовые клетки. Затем проводят инъецирование эмбриональных стволовых клеток в бластоцисты, которые затем переносят псевдобеременным самкам мышей. В заключение проводят получение гомозиготных по разрушенному аллелю гена CRFR2. Кроме того, предложен способ скрининга соединений, которые контролируют кровяное давление через путь рецептора 2 кортиколиберина, с использованием полученных трансгенных мышей. Предложенная группа изобретений позволяет получать трансгенных мышей, которые не экспрессируют белок рецептора 2 кортиколиберина ни по одному аллелю. Изобретение может быть использовано в нейробиологии, эндокринологии и психиатрии для изучения тревожности и депрессии, а также для получения средств для лечения этих заболеваний. 2 с. и 2 з.п. ф-лы, 6 ил. Предпосылки изобретения Участие Федерального финансирования Настоящее изобретение получено частично с использованием фондов федерального правительства по грантам № NIH DK-26741 и NRSA совместно с DK09841 и DK09551. Соответственно, федеральное правительство имеет определенные права на это изобретение. Область изобретения Настоящее изобретение относится к области нейробиологии, эндокринологии и психиатрии. Более конкретно настоящее изобретение относится к изучению тревожности и мышей с дефицитом рецепторов 2 кортиколиберина. Описание уровня техники Кортиколиберин (CRF) является основным координатором гипоталамо-гипофизарно-надпочечниковой системы (НРА). В ответ на воздействие стресса кортиколиберин, высвобождаемый из паравентрикулярных ядер гипоталамуса (PVN), активирует рецепторы кортиколиберина на передних кортикотропах гипофиза, что приводит к выделению в поток крови адренокортикотропного гормона (АСТН). АСТН в свою очередь активирует рецепторы АСТН в корковом веществе надпочечников, что приводит к увеличению синтеза и высвобождению глюкокортикоидов [1]. Рецепторы CRF, CRFR1 и CRFR2 расположены во всей ЦНС и на периферии. Хотя CRF обладает более высоким сродством к CRFR1, нежели к CRFR2, считают, что урокортин (UCN), считается, что пептид, родственный к CRF, является эндогенным лигандом для CRFR2, так как он связывается с почти 40-кратной более высокой афинностью, чем CRF [2]. CRFR1 и CRFR2 обладают примерно 71% сходством по аминокислотной последовательности и отличаются по своей локализации в тканях мозга и в периферических тканях [3-6]. CRFR1 экспрессируется, главным образом в гипофизе, коре, мозжечке, ромбовидном мозге и обонятельной луковице, тогда как CRFR2 находится в латеральной перегородке, вентрально-медиальном гипоталамусе (VMH), хориоидальном сплетении и во многих других периферических участках [5, 7, 8]. CRFR2 имеет несколько изоформ, одна из которых, как было показано, не связывает ни один из известных лигандов [9]. Мыши с дефицитом CRFR1 отличаются пониженными уровнями гормонов НРА системы, ослабленной реакцией на стресс и ослабленным тревожным поведением [10, 11]. Эти результаты совпадают с результатами, полученными с использованием CRFR1 специфических антагонистов in vivo [12-14]. Напротив, CRFR2 специфические антагонисты в настоящее время недоступны, и с момента их клонирования в 1995 очень мало было выяснено относительно физиологических функций CRFR2. UCN может быть эндогенным лигандом для CRFR2, и было показано, что он является модулятором питания при центральном введении [15]. Так как CRFR2 локализован в вентрально-медиальном гипоталамусе, центральном участке регуляции потребления пищи и безопасности, возможно, что действие урокортина на эти рецепторы может влиять на питание. Кроме того, периферическое введение урокортина приводит к гипотонии [2, 16], что может быть результатом действия CRFR2 в сосудистой сети [5, 8]. Предшествующий уровень техники располагает скудными сведениями о мышах с дефицитом рецепторов 2 кортиколиберина. Настоящее изобретение восполняет эту давнишнюю необходимость и потребность. Краткое содержание изобретения Для выяснения развития и физиологической роли CRFR2 были созданы и проанализированы CRFR2 ноль мутантные мыши. Мыши с дефицитом CRFR2 демонстрируют усиленное похожее на тревогу поведение и гиперчувствительную НРА систему в ответ на воздействие стресса. CRFR1 и CRFR2 мутантные мыши представляют ценные модели тревожности и депрессии, и могут, кроме того, помочь в выяснении молекулярного механизма, лежащего в основе этих заболеваний. Изучение сигнальной схемы кортиколиберина и его роли в развитии тревожности и депрессии может дать необходимые подсказки, которые нужны для эффективного лечения этих заболеваний. Таким образом, настоящее изобретение направлено на неприродных трансгенных мышей с нарушением, по крайней мере, по одному аллелю рецепторов 2 кортиколиберина (CRFR2), таким образом, чтобы указанные мыши не экспрессировали из указанного аллеля белок рецептора 2 кортиколиберина. Предпочтительно, чтобы экзоны 10, 11 и 12 ДНК последовательности указанного аллеля рецептора 2 кортиколиберина были делегированы. У трансгенных мышей эти ДНК последовательности могут быть заменены кассетой гена устойчивости к неомицину. Трансгенная мышь может быть либо гетерозиготной, либо гомозиготной для такой замены. В объем настоящего изобретения включено также потомство, полученное от скрещивания мышей настоящего изобретения и мышей других штаммов. Другим вариантом настоящего изобретения является использование мышей с дефицитом CRFR2 для изучения тревожности или депрессии и для тестирования влияния соединений на тревожность или депрессию. Например, предложен способ скрининга соединений, обладающих модулирующей тревожность активностью, включающий стадии: а) введения указанного соединения трансгенной мыши настоящего изобретения; b) тестирования указанной мыши на тревожное поведение; и с) сравнения похожего на тревогу поведения указанной мыши с похожим на тревогу поведением второй трансгенной мыши настоящего изобретения, которой указанное соединение не вводилось. Кроме того, предложен способ скрининга соединения, обладающего модулирующей депрессию активностью, который включает стадии: а) введения указанного соединения трансгенной мыши настоящего изобретения; b) тестирования указанной мыши на похожее на депрессию поведение; и с) сравнения похожего на депрессию поведения указанной мыши с похожим на депрессию поведением второй трансгенной мыши настоящего изобретения, которой указанное соединение не вводилось. Еще один вариант настоящего изобретения включает использование мышей с дефицитом CRFR2 в аналогичной процедуре скрининга соединений, которые влияют на кровяное давление или ангиогенез. Следующим вариантом настоящего изобретения является использование мышей с дефицитом CRFR2 для изучения физиологии НРА системы, т.е. способ скрининга соединений в отношении их воздействия на ответ гипоталамо-гипофизарно-надпочечниковой системы под воздействием стресса, включающий стадии: а) введения указанного соединения трансгенной мыши настоящего изобретения; b) создания для указанной мыши вызывающей стресс ситуации; с) контроля за уровнями плазматического кортикостерона и адренокортикотропного гормона у указанной мыши; и d) сравнения указанных уровней с соответствующими уровнями у трансгенной мыши настоящего изобретения, которая не находилась в указанной, вызывающей стресс, ситуации. В еще одном варианте настоящего изобретения мышей можно использовать для изучения влияния соединений на ответ НРА системы под воздействием стресса, контролируя уровни плазматического кортикостерона и АСТН. Еще один вариант настоящего изобретения относится к использованию мышей настоящего изобретения при изучении влияния рецепторов 2 кортиколиберина на другие белки, такие как кортиколиберин и урокортин. Дальнейшим вариантом настоящего изобретения является использование мышей с дефицитом CRFR2 для исследования реакций CRFR1 в отсутствие CRFR2. Еще одним вариантом настоящего изобретения является управление активностью CRFR2 для стимуляции или ингибирования васкуляризации. Краткое описание чертежей Для того чтобы стали доступны и понятны в деталях вышеуказанные отличительные особенности, преимущества и цели настоящего изобретения, а также и другие, которые станут более понятны, более конкретные описания настоящего изобретения, кратко суммированные выше, могут быть со ссылкой на некоторые его варианты, которые иллюстрируются прилагаемыми чертежами. Эти чертежи составляют часть настоящего описания. Следует отметить, однако, что прилагаемые чертежи иллюстрируют предпочтительные варианты настоящего изобретения, и поэтому их не следует рассматривать как ограничивающие объем изобретения. Фиг.1А-1Е представляют процедуру, которую используют для создания мышей с дефицитом CRFR2, определения мутантных мышей и их характеристик. Фиг.1А представляет геномную структуру CRFR2 гена, где видны делеции экзонов 10, 11 и 12, которые кодируют половину трансмембранного домена пять до конца трансмембранного домена семь. Представлена также мишеневая конструкция, которую используют для гомологичной рекомбинации. На Фиг.1В представлен разрушенный аллель, определенный с помощью саузернблоттинга хвоста ДНК, выделенной из дикого типа (+/+), гетерозиготных (+/-) и ноль мутантных (-/-) мышей. Фиг.1С представляет результаты авторадиографического связывания 125I-Sauvagine у CRFR2 контрольной (сверху) и мутантной (снизу) мыши. Следует отметить отсутствие CRFR2 связывания у латеральной перегородки CRFR2 мутантной мыши, тогда как CRFR1 кортикальное связывание аналогично связыванию у контрольной мыши. Фиг.1D демонстрирует окрашивание надпочечников гематоксилином и эозином (Н и Е). Не отмечается различий в размерах надпочечников (верхнее изображение) при 10-кратном увеличении или в строении (нижнее изображение) при 20-кратном увеличении С, корковое вещество; М, мозговое вещество; ZG, клубочковая зона, ZF, лучковая зона, ZR, сетчатая зона; n=8. Фиг.1Е демонстрирует Н и Е окрашивание гипофизов, которые были расположены на печени для получения срезов тканей (верхние изображения) при 4-кратном увеличении, n=8. Кортикотропы гипофиза были идентифицированы с помощью анти-АСТН антител [20] (нижние изображения) при 10-кратном увеличении, n=5. P, задняя доля; I, промежуточная доля; А, передняя доля. Не наблюдается заметных анатомических различий для гипофизов или для локализации кортикотропов, или для уровней экспрессии АСТН. Фиг.2А-2D демонстрируют гиперчувствительность НРА системы к воздействию стресса у мутантных животных. * = значительное отличие от контрольных животных дикого типа в те же самые моменты времени р<0,01 при определении с помощью теста Scheffe post-hoc. Плазма получена при безанестезионном ретроорбитальном глазном кровотечении. Фиг.2А демонстрирует предстрессовые уровни АСТН в плазме в 7 утра, n=16. Фиг.2В демонстрирует базальные уровни кортикостерона в плазме в 7 утра и 5 вечера, n=7. Фиг.2С представляет временную зависимость АСТН от стрессовых эффектов при ограничении движений. Фиг.2D демонстрирует тот факт, что уровни кортикостерона в плазме в 7 утра значительно отличаются у ноль мутантных мышей от уровней у контрольных мышей дикого типа в те же самые моменты времени, n=7. Фиг.3А-3В демонстрируют влияние 24-часового голодания на потребление пищи у дикого типа и у мутантных мышей-детенышей одного помета. Фиг.3А демонстрирует основной уровень потребления пищи мутантными мышами (n=7) и уровень после 24-часового периода голодания по сравнению с дикого типа мышами-детенышами одного помета (n=10), р<0,001 в тесте Scheffe post-hoc. Фиг.3В представляет вес дикого типа и мутантных мышей как базальный (не зачерненные прямоугольники) и после 24 часов повторного кормления (зачерненные прямоугольники) после периода голодания. Отмечается отсутствие разницы по весу тела в основной группе или в группе с повторным кормлением. Фиг.4А-4Н демонстрируют усиленное подобное тревожности поведение для мутантных животных в каскадном лабиринте и в тесте открытого пространства. Фиг.4А демонстрирует, что для самцов мышей процент времени, проведенного в открытых кронштейнах (**, р<0,005), и число посещений открытых кронштейнов (*, р<0,02) были значительно меньше для мутантных мышей, нежели для контрольных мышей дикого типа (контрольных n=7, мутантных n=7; среднее ±SEM). Фиг.4В представляет тот же тест, что и представленный на Фиг.4А, но для самок мышей. Процент времени, проведенного в открытых кронштейнах (**, р<0,03), и число посещений открытых кронштейнов (*, р<0,03) было значительно меньше для мутантных мышей, нежели для контрольных мышей дикого типа (контрольных n=9, мутантных n=12; среднее ±3ЕМ). Фиг.4С и 4D демонстрируют, что двигательные активности не отличаются у контрольных и мутантных животных (Фиг.4С, самцы мышей; Фиг.4D, самки мышей) при определении по полному числу посещений закрытых кронштейнов и полному количеству посещений всех кронштейнов. Фиг.4Е демонстрирует отсутствие различий для похожего на тревожность поведения при измерении в эксперименте со светлым/темным ящиком в течение времени, проведенного в светлой части ящика. Фиг.4F демонстрирует отсутствие различий для похожего на тревожность поведения при измерении в эксперименте со светлым/темным ящиком для числа переходов из светлой части в темную часть. Фиг.4G представляет промежуток времени, проведенного во внутренних квадратах устройства открытого пространства (*, р<0,05). Фиг.4Н демонстрирует число пересечений, происходящих во внутренних квадратах (**, р<0,01; контроли, n=5; мутанты, n=7; среднее ±SEM). Фиг.5А-5Е демонстрируют повышенные уровни урокортина и мРНК CRF у мутантных штаммов. Для 5В-5Е все цифры были средними для n=3 для мутантов и мышей дикого типа, ±SEM, *, р<0,05; **, р<0,01; ***, р<0,005, при определении с помощью теста Fischer's PLSD post-hoc. Фиг.5А демонстрирует зерна серебра, образовавшиеся в результате in situ гибридизации для мРНК урокортина в ростральном EW (сверху) при 20-кратном увеличении и мРНК CRF в сAmyg (середина), и параветрикулярных ядрах (внизу) при 10-кратном увеличении. Фиг.5В представляет полуколичественный анализ зерен серебра, использовавшихся для определения количества клеток, экспрессирующих мРНК урокортина в ростральном EW. Фиг.5С представляет среднюю оптическую плотность мРНК урокортина на клетку. Фиг.5D представляет оптическую плотность мРНК CRF в сАmyg. Фиг.5Е представляет оптическую плотность мРНК CRF в паравентрикулярных ядрах. Фиг.6 представляет реакцию сердечно-сосудистой системы на внутривенное вливание 1,0 мкг урокортина для дикого типа (n=5) и мутантных мышей (n=3) (зачерненные прямоугольники). Следует отметить существенно уменьшенную реакцию мутантных мышей на инъекцию урокортина. ***р<0,005. CRPR2 мутантным мышам также вводили вторую инъекцию нитропруссида натрия (0,8 мкг в 100 мкл 0,9% физиологического раствора) с последующим определением артериального давления после UCN вливания (незачерненный прямоугольник). Среднее артериальное давление (MAP) определяют, измеряя кровяное давление. Подробное описание изобретения В соответствии с настоящим изобретением можно использовать известные специалистам обычные методы молекулярной биологии, микробиологии и методики рекомбинантных ДНК. Такие методики полностью описаны в литературе. См., например, Maniatis, Fritsch & Sambrook, "Molecular Cloning: A Laboratory Manual (1982); "DNA Cloning: A Practical Approach," Volumes I and II (D.N. Glover ed. 1985); "Oligonucleotide Synthesis" (M.J. Gait ed. 1984); "Nucleic Acid Hybridization" [B. D. Hames & S.J. Higgins Eds. (1985)]; "Transcription and Translation" [B. D. Hames & S.J. Higgins Eds. (1984)]; "Animal Cell Culture" [R. I. Freshney, ed. (1986)]; "Immobilized Cells And Enzymes" [IRL Press, (1986)]; B. Perbal, "A Practical Guide To Molecular Cloning" (1984). Поэтому появляющиеся в описании термины будут иметь приводимые далее значения. В том смысле, как здесь использован, термин "кДНК" относится к ДНК копии мРНК транскрипта гена. В том смысле, как здесь использован, термин "скрининг библиотеки" относится к способу, в котором используют меченый зонд для определения возможности наличия в соответствующих условиях последовательности, комплементарной зонду, присутствующему в конкретной библиотеке ДНК. Кроме того, "скрининг библиотеки" можно осуществить с помощью ПЦР. В том смысле, как здесь использован, термин "ПЦР" относится к полимеразной цепной реакции, которая составляет сущность патентов США №№4683195 и 4683202, выданных Mullis, а также к другим ее усовершенствованиям, известным в данной области техники. Описываемые здесь аминокислоты должны быть предпочтительно в "L" изомерной форме. Однако любые остатки L-аминокислот могут быть замещены остатками в "D" изомерной форме, если только нужные функциональные свойства связывания иммуноглобулина сохраняются полипептидом. NH2 относится к свободной аминогруппе, расположенной у аминоконца полипептида. СООН относится к свободной карбоксильной группе, расположенной у карбоксильного конца полипептида. Аббревиатуры аминокислотных остатков, известные в данной области, используются для обозначения полипептидов, J Biol. Chem., 243:3552-59 (1969). Следует отметить, что все последовательности аминокислотных остатков представлены здесь формулами, в которых левая и правая ориентация соответствует обычному направлению от аминоконца до карбоксиконца. Кроме того, следует учесть, что горизонтальная черточка в начале или в конце последовательности аминокислотного остатка означает пептидную связь с другой последовательностью одного или более из аминокислотных остатков. Термин "репликон" представляет собой любой генетический элемент (например, плазмиду, хромосому, вирус), который функционирует как автономная единица ДНК репликации in vivo; т.е. способен к репликации под своим собственным контролем. Термин "вектор" представляет собой репликон, такой как плазмида, фаг или космида, к которому может быть присоединен другой ДНК сегмент, с тем, чтобы вызвать репликацию этого присоединенного сегмента. Термин "ДНК молекула" относится к полимерной форме дезоксирибонуклеотидов (аденина, гуанина, тимина или цитозина) либо в одноцепочечной форме, либо в форме двухцепочечной спирали. Этот термин относится только к первичной или вторичной структуре молекулы и не ограничивает ее любые конкретные третичные формы. Таким образом, этот термин включает двухцепочечные ДНК, находящиеся inter alia, в линейных молекулах ДНК (например, в рестрикционных фрагментах), вирусах, плазмидах и хромосомах. При обсуждении структуры здесь используют в соответствии с общепринятым только последовательности в направлении 5' Термин "источник репликации" относится к тем ДНК последовательностям, которые участвуют в синтезе ДНК. Термин "кодирующая последовательность" ДНК относится к двухцепочечной ДНК последовательности, которая транскрибируется и транслируется в полипептид in vivo, будучи помещена под контроль соответствующих регуляторных последовательностей. Границы кодирующей последовательности определяются стартовым кодоном у 5' (амино) конца и “стоп”-кодоном трансляции у 3' (карбоксильного) конца. Кодирующая последовательность может включать (но этим не ограничивается) прокариотические последовательности, кДНК из эукариотических мРНК, геномные ДНК последовательности из ДНК эукариот (например, млекопитающих) и даже синтетические ДНК последовательности. Сигнал полиаденилирования и последовательность терминации транскрипции обычно расположены в направлении 3' к кодирующей последовательности. Контрольные последовательности транскрипции и трансляции являются ДНК регуляторными последовательностями, такими как промоторы, энхансеры, сигналы полиаденилирования, терминаторы и т.д., которые обеспечивают экспрессию кодирующей последовательности в клетке хозяина. "Промоторная последовательность" является регуляторным участком ДНК, способным связывать РНК полимеразу в клетке и инициировать транскрипцию в прямом направлении (направление 3') кодирующей последовательности. Для целей определения настоящего изобретения промоторная последовательность связана по своему 3' концу с сайтом инициации транскрипции и простирается в обратном направлении (направление 5'), включая минимальное число оснований или элементов, необходимых для инициации транскрипции на уровнях, детектируемых по отношению к фону. Внутри промоторной последовательности находится сайт инициации транскрипции, а также домены связывания белка (консенсусные последовательности), которые ответственны за связывание РНК полимеразы. Эукариотические промоторы часто, но не всегда, содержат боксы "ТАТА" и боксы "CAT". Прокариотические промоторы содержат последовательности Shine-Dalgarno в дополнении к -10 и -35 консенсусным последовательностям. "Контролирующая экспрессию последовательность" является ДНК последовательностью, которая контролирует и регулирует транскрипцию и трансляцию другой ДНК последовательности. Кодирующая последовательность находится "под контролем" контрольных последовательностей транскрипции и трансляции в клетке, когда РНК полимераза транскрибирует кодирующую последовательность в мРНК, которая затем транслируется в белок, кодируемый кодирующей последовательностью. "Сигнальная последовательность" может быть включена вблизи кодирующей последовательности. Эта последовательность кодирует N-конец сигнального пептида до полипептида, который соединяется с клеткой-хозяином для направления полипептида к поверхности клетки или для секреции полипептида в среду, и этот сигнальный пептид отщепляется клеткой-хозяином до того, как белок выходит из клетки. Сигнальные последовательности можно обнаружить связанными с различными белками, нативными для прокариот и эукариот. Термин "олигонуклеотид" в том смысле, как здесь использован, при упоминании зондов настоящего изобретения, определен как молекула, состоящая из двух или более рибонуклеотидов, предпочтительно, более трех. Его точный размер зависит от многих факторов, которые, в свою очередь, зависят от конечных функций и использования этого олигонуклеотида. Термин "праймер" в том смысле, как здесь использован, относится к олигонуклеотиду, независимо от того, существует ли он в природе в виде очищенного продукта рестрикционного расщепления или получен синтетически, который способен действовать как центр инициации синтеза, будучи помещен в условия, в которых индуцируется синтез продукта удлинения праймера, который комплементарен цепочке нуклеиновой кислоты, т.е. в присутствии нуклеотидов и индуцирующего агента, такого как ДНК полимераза, и при подходящей температуре и рН. Праймер может быть либо одноцепочечным, либо двухцепочечным и должен быть достаточно длинным, чтобы служить затравкой для синтеза целевого продукта удлинения в присутствии индуцирующего агента. Конкретная длина праймера будет зависеть от многих факторов, включая температуру, источник праймера и используемый метод. Например, для диагностических целей, в зависимости от сложности мишеневой последовательности, олигонуклеотидный праймер обычно содержит 15-25 или более нуклеотидов, хотя он может содержать и меньшее число нуклеотидов. Здесь праймеры выбирают таким образом, чтобы они были по существу комплементарны различным цепочкам конкретной мишеневой ДНК последовательности. Это означает, что праймеры должны быть достаточно комплементарны, чтобы гибридизоваться с соответствующими им цепочками. Поэтому нет необходимости в том, чтобы последовательность праймера отражала точную последовательность матрицы. Например, не комплементарный нуклеотидный фрагмент можно присоединить к 5' концу праймера, причем остальная часть последовательности праймера будет комплементарна цепочке. В другом варианте некомплементарные основания или более длинные последовательности можно включить внутрь праймера, при условии, что последовательность праймера достаточно комплементарна последовательности или гибридизуется с ней и тем самым образует матрицу для синтеза продукта удлинения. В том смысле, как здесь использованы, термины "рестрикционные эндонуклеазы" и "рестрикционные ферменты" относятся к ферментам, каждый из которых разрезает двухцепочечную ДНК у или вблизи конкретной нуклеотидной последовательности. Клетка была "трансформирована" экзогенной или гетерологичной ДНК, если такая ДНК была введена внутрь клетки. Трансформирующая ДНК может или не может быть интегрирована (ковалентно связана) в геном клетки. У прокариот, дрожжей и у клеток млекопитающих, например, трансформирующую ДНК можно включить в эписомальный элемент, такой как плазмида. В отношении эукариотических клеток стабильно трансформированной клеткой является клетка, в которой трансформирующая ДНК интегрируется в хромосому так, что она наследуется дочерними клетками посредством репликации хромосом. Такая стабильность демонстрируется способностью эукариотических клеток образовывать клеточные линии или клоны, включающие популяцию дочерних клеток, содержащих трансформирующую ДНК. "Клоном" является популяция клеток, полученных из одной клетки или предшественника в результате митоза. "Клеточной линией" является клон первичных клеток, который способен стабильно расти in vitro в течение многих поколений. Обычно векторы экспрессии, содержащие промоторные последовательности, которые облегчают эффективную транскрипцию встроенного ДНК фрагмента, используют в связи с хозяином. Вектор экспрессии обычно содержит точку начала репликации, промотор (промоторы), терминатор (терминаторы), а также специфические гены, которые способны обеспечить фенотипическую селекцию в трансформированных клетках. Трансформированные хозяева можно ферментировать и культивировать в соответствии с известными специалистам способами для достижения оптимального роста клеток. Такие способы, хорошо известные специалистам, можно использовать для конструирования векторов экспрессии, содержащих соответствующие сигналы, регулирующие транскрипцию и трансляцию. См., например, методики, раскрытые в Sambrook et al., 1989, Molecular Cloning: A Laboratory Manual (2nd Ed.), Cold Spring Harbor Press, N.Y. Ген и его последовательности, регулирующие транскрипцию, определяются как "функционально связанные", если последовательности, регулирующие транскрипцию, эффективно регулируют транскрипцию гена. Векторы настоящего изобретения включают, но ими не ограничиваются, плазмидные векторы и вирусные векторы. Настоящее изобретение направлено на мышей с дефицитом CRFR2, которые были созданы для выяснения развития и физиологической роли CRFR2 при тревожности и в схеме НРА системы. Это было осуществлено в результате делении экзонов 10, 11 и 12 в рецепторе 2 кортиколиберина. В настоящем изобретении эти последовательности были заменены кассетой гена устойчивости к неомицину. Мыши могут быть либо гетерозиготными, либо гомозиготными по дефициту CRFR2 и могут быть скрещены с мышами других штаммов. Настоящее изобретение направлено также на использование мышей с дефицитом CRFR2 при изучении тревожности и депрессии, включая способы тестирования соединений в отношении их активности по модулированию тревожности и депрессии. С помощью CRFR2 мышей можно также осуществить скрининг соединений, которые влияют на кровяное давление и ангиогенез. Настоящее изобретение направлено также на использование мышей с дефицитом CRFR2 при изучении молекулярной физиологии гипоталамо-гипофизарно-надпочечниковой системы (НРА). Этих мышей можно использовать для тестирования влияния соединений на реакции НРА системы на стресс. Настоящее изобретение направлено также на использование трансгенных мышей при изучении влияния молекулярных функций рецепторов 2 кортиколиберина на кортиколиберин, рецептор 1 кортиколиберина, урокортин и другие рецепторы CRF и урокортина. Кроме того, настоящее изобретение можно использовать для изучения реакций и активностей CRFR1 в среде, не содержащей CRFR2. Таким образом, можно изучать CRFR1 реакции без модуляции CRFR2. Настоящее изобретение направлено также на использование агонистов или антагонистов CRFR2 активности для стимуляции или ингибирования васкуляризации. Использование агонистов для стимуляции CRFR2 активности может быть полезным для ингибирования васкуляризации при лечении рака. Приводимые далее примеры представлены для иллюстрации различных вариантов изобретения и никоим образом не ограничивают настоящего изобретения. Пример 1 Создание мышей с дефицитом CRFR2 Для конструирования CRFR2 ноль мутантных мышей из геномной библиотеки ДНК мышиного штамма 129 выделяют геномный клон ДНК, содержащий CRFR2 участок. Из этого клона конструируют направляющий вектор, в котором экзоны 10, 11 и 12 CRFR2 гена, кодирующего участок с начала пятнадцатого трансмембранного домена до конца семнадцатого трансмембранного домена, заменены кассетой гена устойчивости к неомицину (Фиг.1А). Полученную плазмидную ДНК линеаризуют NotI, и осуществляют электропорацию в Jl эмбриональные стеблевые (ES) клетки указанным ранее способом [10]. После селекции в 0,2 мг/мл G418 (активная форма) в течение 7-9 дней устойчивые к неомицину клоны отдельно селектируют и скринируют на присутствие разрушенного CRFR2 аллеля с помощью саузернблоттинга. Позитивные ES клоны инъецируют в C57BL/6 бластоциты для получения химерных мышей. Химерных самцов скрещивают с химерными самками, получая C57BL/6-129 смешанный фенотип мышей. Передачу зародышевой линии разрушенного аллеля определяют с помощью саузернблоттинга ДНК хвоста, полученной из F1 мышат со шкуркой цвета агути (Фиг.1В). Пример 2 Анализ экспрессии CRFR1 и CRFR2 у мышей с дефицитом CRFR2 Для того чтобы определить, привела ли направленная делеция к ноль мутации CRFR2 гена, осуществляют рецепторную авторадиографию на срезах мозга дикого типа контрольных и мутантных животных. Препараты, содержащие 20 мкм срезы тканей мозга, оттаивают при комнатной температуре и дважды промывают в течение 10 минут в 50 мМ Tris буфере (рН 7,4) при комнатной температуре. Затем срезы инкубируют в буфере, содержащем 50 мМ Tris (рН 7,4), 125I-Sauvagine, 10 мМ MgCl2, 0,1% BSA и 0,05% бацитрацина в течение 60 минут при комнатной температуре. Sauvagine представляет собой CRF-родственный пептид и является агонистом CRF рецепторов. Неспецифическое связывание определяют в прилегающих срезах, которые экспонируют как 125I-Sauvagine, так и 1 мкм холодному sauvagine. После окончания периода инкубировании препараты дважды промывают в 50 мМ Tris буфере плюс 0,01% Triton X-100 при 4°С по 5 минут каждым. Препараты быстро погружают в деионизированную воду, сушат и помещают на пленку на 3 дня. У мутантных мышей не обнаруживается связывания в участках мозга, специфичных для CRFR2 (латеральная перегородка), но связывание с CRFR1 в коре сохраняется (Фиг.1С). Эти результаты показывают, что разрушение CRFR2 гена в результате приводит к ноль-мутации у этих мышей. Мутантные мыши были фертильны и передавали мутантный аллель в соответствии с законом Менделя. Пример 3 Гистологический анализ мышей с дефицитом CRFR2 Для определения того, является ли развитие НРА системы угрожающим для мышей с дефицитом CRFR2, получают препараты гипофизов и надпочечников самцов мышей в возрасте 10-12 недель и окрашивают их гематоксилином и эозином (Н и Е). Вкратце, мышам вводят 4% параформальдегид (PFA). Ткани удаляют, затем фиксируют в течение ночи при 4°С и осуществляют криозащиту в 30% сахарозе в PBS. Получают препараты тканей толщиной 12 мкм и окрашивают их гематоксилином и эозином. Полученные результаты не выявляют никакой разницы в строении или типах клеток (Фиг.1D-1Е). Кроме того, срезы гипофиза окрашивают анти-АСТН антителами. Получают срезы гипофизов, затем их фиксируют в 4% PFA в течение 5 минут, промывают PBS и окрашивают АСТН антителами, как указано ранее [10]. Не обнаружено качественных различий между дикого типа и мутантными кортикотропами (Фиг.1Е). Пример 4 Определение уровней кортикостерона и АСТН у мышей с дефицитом CRFR2 Для анализов кортикостерона и АСТН плазму получают от содержащихся отдельно мышей в возрасте 10-12 недель. Образцы отбирают из ретроорбитального глазного кровотечения у не анестезированных мышей в течение 30 сек создания беспокойства в клетке. Базальные утренние образцы отбирают в 7.00. Базальные вечерние образцы отбирают в 17.00. В анализе кортикостерона (ICN Biomedicals, Dosta Mesa, CA) используют 5 мкл плазмы, и в АСТН анализе (Nichols Institute Diagnostics, San Juan Capistrano, CA) используют 50 мкл плазмы, измерения проводят дважды с помощью наборов для радиоиммунного анализа. Нормальные базальные уровни АСТН и кортикостерона были обнаружены у мутантных и контрольных животных (Фиг.2А-2В), что соответствует информации, представленной на Фиг.1Е, о том, что уровни АСТН в мозге не изменены. Пример 5 Влияние стресса на реакцию НРА системы у мышей с дефицитом CRFR2 Для исследования реакции НРА системы на стресс животных подвергают стрессу, связанному с физическим ограничением движения, в течение увеличивающегося промежутка времени. Образцы крови отбирают сразу после каждых 2, 5 или 10 минут стресса, связанного с ограничением движения, в 50 мл конические ампулы (пластиковые конические ампулы без дна). У каждой из мышей кровь отбирают только один раз. Образцы плазмы немедленно центрифугируют и хранят при -20°С до проведения анализа. Для анализа кортикостерона (ICN Biomedicals, Costa Mesa, CA) используют 5 мкл плазмы, и для анализа АСТН (Nichols Institute Diagnostics, San Juan Capistrano, CA) используют 50 мкл плазмы, используя наборы для радиоиммунного анализа и осуществляя анализы в двух экземплярах. Уровни АСТН у контрольных животных достигают максимального значения после 10 минут ограничения движения. Напротив, уровни АСТН у животных-мутантов оказываются существенно повышенными и достигают максимального значения только через 2 минуты стресса, вызванного ограничением движения (Фиг.2С). Аналогично, уровни кортикостерона у животных-мутантов оказались существенно повышены после двух минут ограничения движения, тогда как у контрольных животных уровни повышались после пяти минут стресса (Фиг.2С). Эти результаты демонстрируют гиперчувствительную реакцию НРА системы на стресс у мутантных мышей. ПРИМЕР 6 Мыши с дефицитом CRFR2 чувствительны к голоданию Так как CRFR2 находится в избытке в VMH и так как более ранние исследования продемонстрировали аноректическое действие урокортина (15), у мутантных и дикого типа детенышей одного помета определяли основное потребление корма и увеличение веса. Потребление корма определяют для содержащихся отдельно самцов мышей одного помета в возрасте 12-16 недель. Мышей и их гранулированный пищевой рацион взвешивали ежедневно в 9 часов утра. Для эксперимента по голоданию контрольных и мутантных мышей одного помета содержат отдельно и определяют их базовое потребление корма и вес. Мышам не дают корма в течение 24 часов, начиная с 12 часов дня, но потребление воды не ограничивают (ad libidum). После периода голодания мышей взвешивают и дают им заранее определенный кормовой брикет. Затем кормовые брикеты взвешивают каждые два часа до тех пор, пока не гасят свет (18 часов). Кормовые брикеты и мышей снова взвешивают на следующее утро. Регистрируют потерю веса во время голодания, а также полное потребление корма и увеличение веса за 24 часа после периода голодания. Базовое питание и увеличение веса у CRFR2 мутантных (mut) самцов мышей аналогичны результатам для дикого типа (wt) самцов одного помета (24 час базовое потребление корма wt=4,3±0,24 г, mut=4,6±0,23 г; вес тела wt=21,7±0,66 г, mut=21,2±0,50 г; n=10, среднее ± SEM). Для определения того, изменяют ли стрессовые стимулы потребление корма мутантными мышами, контрольных и мутантных мышей лишали корма на 24 часа, а затем снова давали корм, после чего измеряли потребление ими корма и изменения в весе. Результаты, полученные после голодания, свидетельствуют о значительном уменьшении потребления корма мутантными мышами после 24 часов голодания (Фиг.3А). Мутантные мыши потребляют 75% от уровня потребления мышами дикого типа в течение 24 часов после голодания. Однако веса мутантных мышей и мышей дикого типа заметно не отличались после голодания или возобновления кормления (Фиг.3В). Пример 7 Оценка похожего на тревожность поведения у мышей с дефицитом CRFR2 в каскадном лабиринте Так как CRFR1 мутантные мыши демонстрируют анксиолитико-подобное поведение [10], CRFR2 мутантных мышей изучали в аналогичных тестах, используя три различные тестовые схемы. В первой тестовой схеме контрольных и мутантных мышей оценивают, используя каскадный лабиринт (ЕРМ). В этом эксперименте используют самцов и самок мышей в возрасте 22-24 недели. В качестве контроля используют дикого типа мышей одного помета. Животных содержат группами в постоянных условиях темноты/освещенности (свет включают в 6 утра, выключают в 6 вечера) и обрабатывают их через день в течение одной недели до проведения эксперимента. Конструкция каскадного лабиринта выполнена из черного плексигласа и имеет два открытых кронштейна (30×5 см) и два закрытых кронштейна того же размера со стенками высотой 30 см. Эта конструкция приподнята на 30 см над землей. Кронштейны соединены центральной площадкой (5×5 см), и таким образом лабиринт образует знак плюс. 25-ваттная лампа, помещенная сверху, обеспечивает освещенность в 6 люкс для открытых кронштейнов. Все тесты осуществляют во время светлой фазы цикла свет/темнота. Мышам дают привыкнуть к условиям в экспериментальном пространстве в течение 1 часа перед тем, как изучают их поведение, и животных тестируют по отдельности в пятиминутных сеансах. Каждую из мышей помещают в центр платформы мордой к открытому кронштейну для начала тестового сеанса. Оценкой поведения служат количество входов в открытые и закрытые кронштейны и продолжительность времени, проведенного в различных отделах лабиринта. Входы в кронштейны определяют как вход в кронштейн всеми четырьмя лапами. Входы в закрытые кронштейны принимают за показатель двигательной активности в лабиринте. Камера, смонтированная над конструкцией, позволяет наблюдать за поведением животных на видеомониторе, расположенном в соседней комнате. В конце теста количество входов и время, проведенное на открытых кронштейнах, выражают в процентах от полного числа входов в кронштейны и продолжительности теста, соответственно. Результаты выражают как среднее ± стандартная ошибка. Поведенческие параметры, полученные в результате ЕРМ теста, анализируют, используя тест Стьюдента. Полученные результаты показывают, что как самцы, так и самки CRFR2 мутантных мышей проводят меньше времени на открытых кронштейнах и менее часто посещают открытые кронштейны конструкции лабиринта, нежели это делают контрольные мыши дикого типа. Обнаруживается значительный эффект для обоих процентов вхождений в открытые кронштейны для самцов и самок мышей (Фиг.4А и 4В). Усиление похожего на тревожность поведения не связано с изменением двигательной активности, так как полная активность в закрытых кронштейнах и полное количество вхождений в кронштейны не отличаются для двух групп (Фиг.4С и 4D). Эти результаты показывают, что CRFR2 мутантные мыши демонстрируют заметно усиленное похожее на тревожность поведение. Пример 8 Оценка похожего на тревожность поведения у мышей с дефицитом CRFR2 в светлом/темном ящике Поведение CRFR2 мутантных и контрольных мышей анализируют также как похожее на тревожность поведение мышей в светлом/темном ящике. Прямоугольный плексигласовый ящик делят на два отделения, одно из которых выкрашено в белый цвет (28,5 см × 27 см), а другое - в черный цвет (14,5 см × 27,0 см). Интенсивность света составляет 8 люкс в черном отделении, покрытом красной плексигласовой крышкой, и 400 люкс в белом отделении. Отделения соединены отверстием (7,5 см × 7,5 см), расположенным на уровне дна в центре перегородки. Все тесты осуществляют во время темной фазы цикла между 19 и 21 часом. Каждое животное тестируют в течение 10 минут, помещая в центр белой площадки, и регистрируют количество переходов между двумя отделениями и продолжительность времени, проведенного в белом отделении. Камера, смонтированная над конструкцией, позволяет осуществлять наблюдение и регистрацию из соседней комнаты. Результаты, полученные в тесте со светлым/темным ящиком, показывают, что CRFR2 мутантные мыши проводят столько же времени в светлом отделении ящика и осуществляют столько же переходов между светлым и темным отделениями ящика, что и контрольные мыши (Фиг.4Е и 4F). В этом эксперименте не отмечается различий между двумя группами мышей. Пример 9 Оценка похожего на тревожность поведения у мышей с дефицитом CRFR2 в тесте открытого пространства Похожее на тревожность поведение анализируют также для CRFR2 мутантных и контрольных мышей в конструкции открытого пространства. Конструкция открытого пространства представляет собой белый плексигласовый ящик (50×50×22 см) с 16 квадратами (12×12 см), нарисованными на дне (12 внешних и 4 внутренних). Направленная на центр поверхности лампа обеспечивает освещенность дна в 120 люкс. Тестирование осуществляют во время темной фазы цикла свет/темнота в комнате с постоянным фоновым белым шумом (52 децибел). Каждую из мышей помещают в центр конструкции для начала проведения 10-минутного теста. Время (сек), проведенное во внутренних квадратах, брожение (число пересеченных квадратов), дефекацию (число фекальных катышков), подъем на задние лапы и время, потраченное на вылизывание (сек), оценивают по видеозаписи. Пересечения внутренних квадратов выражают как процент от брожения. Результаты теста открытого пространства показывают, что CRFR2 мутантные мыши проводят меньше времени во внутренних квадратах и дают меньший процент пересечений внутренних квадратов, нежели это справедливо для мышей дикого типа (Фиг.4G и Н). Не наблюдается никаких различий в брожении, подъеме на задние лапы, дефекации или вылизывании для различных групп (данные не представлены). Пример 10 Влияние дефицита CRFR2 на экспрессию других генов Так как не было обнаружено никаких значительных анатомических дефектов в компонентах НРА системы (Фиг.1D и 1Е), изменения в стрессовых и поведенческих реакциях у мутантных животных могут быть связаны с изменением генной экспрессии других компонентов CRF сигнальной схемы. Для выяснения этой возможности исследуют экспрессию UCN, CRF и CRFR1 мРНК с помощью in situ гибридизации. In situ гибридизацию осуществляют в соответствии с раскрытыми ранее способами [36]. Вкратце, препараты тканей (20 мкм) фиксируют в 4% параформальдегиде, промывают в PBS, погружают в уксусный ангидрид, дегидратируют с помощью этанола, удаляют липиды в хлороформе и снова дегидратируют. Затем препараты гибридизуют с 35S-меченным рибозондом в 50% деионизированной формамидной гибридизационной смеси в течение ночи при 55°С во влажной инкубационной камере. После инкубирования препараты промывают в IX SSC при комнатной температуре в течение 30 минут при встряхивании, обрабатывают 20 мкг/мл РНКазы (Promega) при 37°С в течение 30 минут, промывают в 1X SSC буфере при комнатной температуре в течение 30 минут, промывают 3Х в течение 20 минут при 65°С в 0,1X SSC при встряхивании, промывают в 0,1X SSC при комнатной температуре в течение 30 минут, дегидратируют с помощью этанола, сушат на воздухе и помещают на гиперпленку Kodak (Eastman Kodak, Rochester, NY) на три дня. После проявления пленки препараты погружают в NTB2 жидкую ядерную эмульсию (Eastman Kodak; разбавлена 1:1 водой), экспонируют в течение 10 дней, фотографически обрабатывают, осуществляют обратное окрашивание гематоксилином и заклеивают покровными стеклами. Препараты анализируют, используя аналитическую систему изображений Image Pro Plus (Media Cybernetics, Silver Springs, MD). Для анализа PVN и cAmyg (площадь = 3022 пикселей) используют циркуль для определения средней оптической плотности для каждого препарата, так что анатомический атлас подобранных срезов для каждого животного сравнивают в идентичных участках PVN и cArayg. EW клеточные тельца, экспрессирующие урокортин, оказываются слишком диффузными, чтобы их проанализировать, используя стандартные способы измерения оптической плотности. Поэтому используют такие параметры, чтобы компьютер определял число клеток внутри указанных EW, отражающих минимальную оптическую плотность, по цвету и размеру клеток, как определено заранее, чтобы исключить не позитивные клетки и фоновые зерна серебра. Каждая клетка, которая по компьютерным данным рассматривается как позитивная для мРНК урокортина, затем измеряется по оптической плотности. Затем сравнивают среднюю оптическую плотность и количество клеток для каждого среза. Как видно на Фиг.5А, мРНК урокортина значительно увеличена в ростральном участке Edinger Westphal (EW) ядер как для количества экспрессирующих клеток (Фиг.5В), так и для плотности мРНК урокортина на клетку (Фиг.5С) у мутантных животных. Центральные ядра миндалины (cAmyg) демонстрируют значительное увеличение CRF мРНК для ноль мутантных животных (Фиг.5А и 5D). У контрольных, не подвергавшихся стрессу животных, не обнаружено заметных изменений в мРНК CRF в PVN (Фиг.5А и 5Е). Характер экспрессии или уровни мРНК CRFR1 в мозге или передней доле гипофиза не отличаются для мутантных и дикого типа мышей (данные не представлены). Эти результаты показывают, что CRFR2 мутантные мыши отличаются повышенными уровнями экспрессии мРНК CRF в cAmyg и мРНК урокортина в ростральных Edinger Westphai ядрах. Пример 11 Оценка гипотонии в ответ на UCN у CRFR2 мутантных мышей В более ранних сообщениях указывалось на гипотоническую реакцию в ответ на периферическое введение урокортина [2]. Кроме того, CRFR2 были локализованы в эндотелиальных клетках сосудов [5, 8], и была выдвинута гипотеза о том, что они ответственны за вазодилататорную активность урокортина. Для проверки этой гипотезы CRFR2 мутантным и контрольным мышам вводят инъекции урокортина и измеряют изменения их кровяного давления. Исследуют у анестезированных изофлорином мышей реакции сердечно-сосудистой системы на внутривенные вливания урокортина и нитропруссида натрия, вазодилататора (дикий тип: n=5; мутанты: n=3). Артериальный катетер для регистрации кровяного давления изготовлен из стерильной РЕ-10 трубки, размягченной и вытянутой до внешнего диаметра примерно 0,4 мм. Высвобождают бедренную артерию, и артериальный катетер, заполненный гепариновым солевым раствором (500 ед/мл), имплантируют и фиксируют хирургическими нитками и клеем для тканей (Vetbond). Катетер соединяют с датчиком кровяного давления (Statham), и импульсы кровяного давления подают на Gould самописец. Затем имплантируют второй катетер во внешнюю яремную вену для внутривенного введения лекарств. Вливание лекарства осуществляют через 30 минут после завершения процедуры введения канюль. Венозный катетер соединяют с заполненным лекарством шприцом. Вливание завершают за 0,5-1,0 мин. Мышам дикого типа и мутантным мышам вводят идентичные дозы урокортина (0,1 мкг в 200 мкл 0,9% физиологического раствора) и физиологический раствор (в качестве контроля). Используемые дозы определяют в предварительных экспериментах с использованием результатов, полученных в соответствующих исследованиях на крысах штамма Sprague Dawley [2]. Для подтверждения того, что отсутствие реакции сердечнососудистой системы на инъекции урокортина у мутантных животных не связано с потерей у мышей способности к вазодилятации, мутантным мышам вводят также в качестве второго вливания нитропруссид натрия (0,8 мкг в 100 мкл 0,9% физиологического раствора) после восстановления артериального давления после вливания урокортина. Среднее артериальное давление (MAP) определяют на основании записей кровяного давления. Внутривенное вливание урокортина (0,1 мкг) приводит к заметной депрессорной реакции (-28,3±2,0 мм рт. ст.) у контрольных мышей (Фиг.6). Снижение артериального давления наблюдается на протяжении периода записи (90-120 мин). Совершенно иначе, у мутантных мышей не наблюдается измеряемой реакции на урокортин (только 1 из исследованных мутантных мышей проявила очень маленькое и кратковременное снижение артериального давления (-3,5 мм рт. ст.), что, по-видимому, связано с самим давлением в процессе инъекции (Фиг.6)). Для подтверждения того, что периферическая сосудистая система мутантных мышей способна к вазодилятации в ответ на другие стимулы, мутантным мышам вводят нитропруссид натрия (NP), который вызывает вазодилятацию как донор окиси азота. Быстрая и существенная депрессорная реакция постоянно наблюдается в ответ на инъекции ниропруссида натрия (-30,0±5,0 мм рт. ст.), как видно на Фиг.6 (белый прямоугольник). Базовое MAP под анестезией незначительно отличается у мутантных мышей (76 мм рт. ст.) и дикого типа мышей (74 мм рт. ст.) во время этого эксперимента. Пример 12 CRFR2 влияет на васкуляризацию Гистологический анализ органов CRFR2 мутантных мышей выявил, что васкуляризация некоторых органов, включая мозг, гипофиз и надпочечники, заметно выше у мышей с дефицитом CRFR (данные не представлены). Это указывает на то, что CRFR2 могут действовать как непосредственные регуляторы васкуляризации. Таким образом, при болезненных состояниях для управления васкуляризацией можно использовать лиганды, служащие агонистами или антагонистами CRFR2. Одним из примеров является рак, когда усиленная васкуляризация необходима для роста раковых клеток. Стимулируя активность CRFR2, агонист можно использовать для ингибирования васкуляризации и тем самым затруднить рост раковых клеток. Пример 13 Суммарные эффекты CRFR2 делеций на тревожность и стресс Представленные здесь результаты предполагают, что CRFR2 ноль мутантные мыши проявляют стрессо-чувствительный и похожий на тревожность фенотип. Хотя базовое кормление и увеличение веса были нормальными, мутантные мыши реагировали на голодание, потребляя меньшее количество корма в период восстановления кормления после стресса, вызванного голоданием. Хотя это может быть связано с метаболизмом, так как мутантные и контрольные животные не демонстрируют никакой разницы в увеличении или снижении веса во время эксперимента, возможно, что стресс в результате голодания изменяет состояние тревожности у животных, тем самым снижая их аппетит или влияя на их метаболизм. Мутантные мыши демонстрируют также быструю НРА реакцию на стресс, связанный с ограничением движения, что снова дает возможность предположить, что эти животные более чувствительны к стрессу. Снижение уровней АСТН у мутантов, которое наблюдается после 10 минут ограничения, может быть результатом более быстрого негативного обратного воздействия глюкокортикоидов на гипоталамус, так как мутантные мыши демонстрируют более высокие уровни стероидов раньше, чем контрольные мыши. Поэтому нельзя отбросить возможность второго механизма, приводящего к активации надпочечников у мутантных мышей. Взятые вместе результаты кормления и НРА системы предполагают наличие гиперчувствительности к стрессу у CRFR2 мутантных мышей, хотя другие физиологические объяснения могут быть включены в реакции на изменения питания или повышения скорости, с которой НРА системы мутантных мышей реагирует на стресс. Мутантные мыши демонстрируют также повышенное похожее на тревожность поведение в тесте ЕРМ и в тесте открытого пространства. Однако эти мыши демонстрируют аналогичные уровни похожего на тревожность поведения в светлом/темном ящике. Хотя фармакологическая чувствительность и специфичность были продемонстрированы во многих тестах на тревожность у животных, иногда наблюдаются различия в выполнении заданий [17, 18]. Поведение в системе светлое/темное может отличаться в том, что эта задача более коррелирует с реакциями на новизну (неофобия), нежели с исследованиями [19], хотя поведение в ЕРМ определяется исследованием создающего беспокойство окружения [19]. Условия освещенности во время тестирования также могут оказать значительное влияние на способность определять анксиолитические или анксиогенные эффекты у тестируемых животных [17]. Такой характер результатов, полученных для CRFR2 мутантных мышей, демонстрирует повышенную эмоциональность, связанную с исследованием создающего беспокойство окружения, но не неофобию. Результаты, полученные для CRFR2 мутантных мышей, предполагают, что эти различия в поведенческих тестах можно объяснить различиями в похожем на тревожность поведении, обнаруженном у мышей. Пример 14 Возможные воздействия на тревожность повышенных CRF в сАmyg Повышенные мРНК CRF в сАmyg могут объяснить похожее на тревожность поведение и повышенную чувствительность НРА системы мутантных мышей, так как эти ядра экспрессируют CRFR1 [8] и играют главную роль в передаче стрессовых сигналов [20]. Кроме того, было показано, что перегородка, которая содержит CRFR2 в избытке, модулирует активность миндалины [21-23], и поражение этих ядер приводит к пониженной секреции АСТН после стресса, связанного с ограничением движения [24-27]. Было показано, что поражения миндалины блокируют индуцированную CRF тревожность [20], а также гиперэмоциональность, вызываемую поражениями перегородки [21]. Такая нервная схема может объяснить ослабленное похожее на тревожность поведение, которое наблюдается у мышей с дефицитом CRFR1 [10], так же, как усиленное похожее на тревожность поведение у мышей с дефицитом CRFR2. Поэтому CRFR2 мутантные мыши представляют возможное доказательство нового механизма рецепторной модуляции похожего на тревожность поведения. Возможно, что во время стресса CRFR2 в латеральной перегородке модулирует активность миндалины, и в отсутствии CRFR2 свободная активность миндалины может привести к усиленному похожему на тревожность поведению. CRFR2 в латеральной перегородке может функционировать как ингибитор воздействия PVN на НРА реакции на стресс. Так как у мутантных мышей наблюдается дефицит CRFR2 в латеральной перегородке, вызванная стрессом активация PVN может происходить более быстро. Кроме того, CRFR2 является преобладающим CRF рецептором, экспрессируемым PVN у не подвергавшихся стрессу животных, тогда как CRFR1 обнаруживается в PVN только в условиях стресса [28, 29]. Поэтому при отсутствии CRFR2 во время стресса локальные эффекты на активность PVN могут быть изменены. Пример 15 Возможные механизмы тревожности, вызываемые повышенным содержанием мРНК UCN в ростральных EW Повышение мРНК урокортина в ростральных EW может быть вторым механизмом, приводящим к повышенному похожему на тревожность поведению мутантных мышей, так как было показано, что урокортин вызывает похожее на тревожность поведение, будучи введенным внутривенно [30]. Дополнительные объяснения повышенного похожего на тревожность поведения, такого как повышенная чувствительность автономной нервной системы [31-33], также до сих пор нельзя было отбросить. Более ранние исследования с использованием антисмысловых олигонуклеотидов обнаружили противоречивые результаты относительно роли CRFR2 в тревожности и поведении, хотя эти сообщения не продемонстрировали эффекта, похожего на анксиолитический, при инъекциях CRFR1 антисмысловых олигонуклеотидов [34, 35]. Пример 16 CRFR2 ноль мыши и чувствительность автономной нервной системы Дополнительные объяснения повышенного похожего на тревожность поведения, такого как повышенная чувствительность автономной нервной системы [31-33], также нельзя отбросить. Более ранние исследования с использованием антисмысловых олигонуклеотидов обнаружили противоречивые результаты относительно роли CRFR2 в тревожности и поведении [34, 35]. Хотя эти сообщения продемонстрировали похожий на анксиолитический эффект при инъекциях CRFR1 антисмысловых олигонуклеотидов, ни в одном из этих исследований нет сообщений о четких данных относительно инъекций CRFR2 антисмысловых олигонуклеотидов. Хотя методика инъекций антисмысловых олигонуклеотидов предлагает возможные перспективы, она остается предметом тщательного изучения, так как полное исключение мишени не может быть заменено пониженными уровнями белка, что имеет место у нокаутированных животных. Пример 17 Влияние UCN на вазодилятацию Отсутствие CRFR2 у ноль мутантных мышей дает возможность подтвердить воздействие урокортина на вазодилятацию. У мутантных мышей отсутствует реакция на внутривенное введение урокортина, хотя животные дикого типа демонстрируют резкое понижение среднего артериального давления. Инъекция нитропруссида приводит к вазодилятации у мутантных мышей, подтверждая тот факт, что отсутствие реакции на урокортин не связано с физической неспособностью сосудистой системы мутантов к расширению, но специфически связано с отсутствием CRFR2. Эти результаты подтверждают гипотезу о том, что воздействие урокортина на гипотонию [2, 16] осуществляется за счет действия CRFR2 в эндотелиальных клетках сосудов [5, 8], так как CRFR2 мутантные мыши не реагируют на урокортин. Хотя в настоящее время не известны физиологические стимулы, под действием которых наиболее вероятно осуществляется вызываемая UCN вазодилятация, воздействие урокортина на CRFR2 в сосудистой системе может оказаться интересным фактом с точки зрения создания лекарств для лечения гипертонии. Выводы В итоге эти результаты показывают, что мыши с дефицитом CRFR2 демонстрируют повышенное похожее на тревожность поведение и гиперчувствительность НРА системы при реакции на стресс. CRFR1 и CRFR2 мутантные мыши представляют собой полезные модели для изучения тревожности и депрессии и могут в дальнейшем помочь определить молекулярные механизмы, лежащие в основе этих заболеваний. Изучение CRF сигнальной схемы и ее роли в управлении тревожностью и депрессией может предоставить необходимые ключи для эффективного лечения этих заболеваний. Далее перечислены используемые в описании ссылки на источники информации. 1. Vale, W., Spiess, J., Rivier, C. & Rivier, J., Science 213, 1394-1397 (1981). 2. Vaughan, J., et al., Nature 378, 287-292 (1995). 3. Kishimoto, Т., Pearse, R.V., 2nd, Lin, C.R. & Rosenfeld, M.G., Proc. Nati. Acad. Sci. USA 92, 1108-1112 (1995). 4. Lovenberg, T.W., et al., Proc. Natl. Acad. Sci. USA 92, 836-840 (1995). 5. Perrin, M., et al., Proc. Natl. Acad. Sci. USA 92, 2969-2973 (1995). 6. Stenzel, P., et al., Mol. Endocrinol. 9, 637-645 (1995). 7. Potter, E., et al., Proc. Natl. Acad. Sci. USA 91, 8777-8781 (1994). 8. Chalmers. D.T., Lovenberg, T.W. & De Souza, E.B, J. Neurosci. 15, 6340-6350 (1995). 9. Miyata, I., et al., Biochem. Biophys. Res. Conunun. 256, 692-696 (1999). 10. Smith, G.W., et al., Neuron 20, 1093-102 (1998). 11. Tirnpl, P., et al., Nat. Genet. 19, 162-166 (1998). 12. Webster, E.L., et al., Endocrinology 137, 5747-50 (1996). 13. Bornstein, S.R., et al., Endocrinology 139, 1546-55 (1998). 14. Deak, Т., et al., Endocrinology 140, 79-86 (1999). 15. Spina, M., et al., Science 273, 1561-4 (1996). 16. Schilling, L., Kanzler, С., Schmiedek, P. & Ehrenreich, H., Br. J. Pharmacol. 125, 1164-1171 (1998). 17. Hogg, S., Pharmacol. Biochem. Behav. 54, 21-30 (1996). 18. Rodgers, R.J, Benav. Pharmacol. 8, 477-496 (1997). 19. Belzung, С. & Le Pape, G., Pnysiol. Benav, 56, 623-628 (1994). 20. Liang, К.С., et al., J. Neurosci. 12, 2313-2320 (1992). 21. King, F.A. & Meyer, P.M. Science 128, 655-656 (1958). 22. Melia, K.R. & Davis, M, Physiol. Benav. 49, 603-611 (1991). 23. Lee, Y. & Davis, M, J. Neurosci. 11, 6424-6433 (1997). 24. Alien, J. & Alien, C, Neuroendocrinology 19, 115-25 (1975). 25. Beaulieu, S., Di Paolo, Т. & Barden, N., Neuroendocrinology 44, 247-54 (1986). 26. Beaulieu, S., Di Paolo, Т., Cote, J. & Barden, N., Neuroendocrinology 45, 37-46 (1987). 27. Marcilhac, A. & Siaud, P, Exp. Physiol. 81, 1035-1038 (1996). 28. Imaki, et al., Brain Res. Mol. Brain Res. 38, 166-170 (1996). 29. Imaki, Т., et al., Endocr. J. 43, 629-638 (1996). 30. Moreau, J.L., Kilpatrick, G. & Jenck, F, Neuroreport 8, 1697-701 (1997). 31. Udelsman, R., et al., Nature 319, 147-50 (1986). 32. Andreis, P.G., Neri, G. & Nussdorfer, G.G., Endocrinology 128, 1198-200 (1991). 33. Andreis, P.G., Neri, G., Mazzocchi, G., Musajo, F. & Nussdorfer, G.G., Endocrinology 131, 69-72 (1992). 34. Heinrichs, S.C., Lapsansky, J., Lovenberg, T.W., De Souza, E.B. & Chalmers, D.T., Regul Pept. 71, 15-21 (1997). 35. Liebsch, G., Landgraf, R., Engelmann, M., Lorscher, P. & Holsboer, F., [In Process Citation] J. Psychiatr. Res. 33, 153-163 (1999). 36. Bale, T.L. & Dorsa, D.M., Endocrinology 136, 27-32 (1995). Любые патенты или публикации, упомянутые в этом описании, являются показателями уровней знаний специалистов в области, к которой относится настоящее изобретение. Эти патенты и публикации включены сюда для ссылки в той же степени, как если бы каждая отдельная публикация была конкретно и индивидуально включена сюда для ссылки. Специалистам легко понять, что настоящее изобретение адаптировано для выполнения целей и достижения указанных результатов и преимуществ, а также тех, которые ему присущи. Представленные примеры, наряду с описанными здесь способами, процедурами, обработками, молекулами и конкретными соединениями, являются просто представителями предпочтительных вариантов, являются примерами и не должны ограничивать объем настоящего изобретения. Изменения в них и другие применения будут очевидны специалистам, но все они включены в объем настоящего изобретения, определяемый формулой изобретения. Формула изобретения1. Способ получения неприродной трансгенной мыши гомозиготной по разрушенным аллелям эндогенного гена рецептора 2 кортиколиберина (CRFR2), при этом указанная мышь не экспрессирует белок рецептора 2 кортиколиберина ни по одному аллелю, включающий следующие стадии: a) выделение геномного клона, содержащего локус гена CRFR2; b) конструирование направляющего вектора, в котором экзоны 10, 11 и 12 гена CRFR2 заменены кассетой селективного маркерного гена; c) перенос указанного вектора в эмбриональные стволовые клетки; и d) инъецирование указанных эмбриональных стволовых клеток в бластоцисты, которые затем переносят псевдобеременным самкам мышей для воспроизведения мышей, гомозиготных по разрушенным аллелям гена CRFR2. 2. Способ по п.1, отличающийся тем, что кассетой селективного маркерного гена является кассета гена устойчивости к неомицину. 3. Способ по п.1, дополнительно включающий следующие стадии: a) скрещивание мышей, полученных в соответствии со способом по п.1, с мышами другого штамма; и b) селекция потомства, полученного в результате указанного скрещивания, по гомозиготности по разрушенным аллелям гена CRFR2. 4. Способ скрининга соединений, которые контролируют кровяное давление через путь рецептора 2 кортиколиберина, включающий следующие стадии: a) введение соединения трансгенной мыши, полученной способом по п.1; b) тестирование указанной трансгенной мыши по изменениям кровяного давления; и c) сравнение изменений кровяного давления у указанной трансгенной мыши с изменениями кровяного давления у мыши дикого типа, которой указанное соединение также вводили, при этом отсутствие изменений кровяного давления в указанной трансгенной мыши свидетельствует о том, что указанное соединение контролирует кровяное давление через путь рецептора 2 кортиколиберина. MM4A - Досрочное прекращение действия патента СССР или патента Российской Федерации на изобретение из-за неуплаты в установленный срок пошлины за поддержание патента в силе Дата прекращения действия патента: 15.07.2006 Извещение опубликовано: 20.12.2007 БИ: 35/2007 Популярные патенты: 2407282 Способ выращивания корнесобственных саженцев винограда и машина для его осуществления ... черенки с двумя-тремя почками с шагом высадки 0,12 0,13 м в два ряда вдоль гибких поливных трубопроводов. А при температуре почвы выше +35°С в пленке с шагом 1 м делают локальные разрезы поперек гребня шириной 0,6 0,8 м. При температуре воздуха выше +40°С на светопроницаемую пленку опрыскиванием наносят побелку. Машина для формирования гребней для выращивания корнесобственных саженцев винограда содержит раму, кронштейны навески на тяги агрегатируемого трактора и опорные колеса. Машина снабжена последовательно установленными на раме с возможностью переустановки по глубине обработки и ширине захвата стойками рыхлителей, сферическими дисковыми рабочими органами, левым и ... 2051971 Способ определения биологической активности -эндотоксинов различных патотипов bacileus thuringiensis ... буфером рН 7,5 и используют для определения антибактериальной активности. В таблице приводятся данные, полученные при усреднении значений для 3 концентраций растворов в 3 повторностях. П р и м е р 2. Берут суспензию спор и кристаллов B.thuringiensis subsp. tenebrionis, полученную так, как описано в примере 1, промывают так, как описано в примере 1, растворяют (без отделения спор) в 0,035 н.NaOH так, как описано в примере 1. -эндотоксинов различных патотипов bacileus thuringiensis, патент № 2051971" SRC="/images/patents/421/2051145/948.gif" > -Эндотоксины осаждают из супернатанта уксусной кислотой при рН 6,5, центрифугируют на холоду при 15 тыс.об./мин, 20 мин, осадок ... 2206985 Упряжь для собак ... шлейки выполнены в виде многослойных ремней из эластичного материала, между слоями спинного ремня размещена вставка из синтетического материала, а в месте соединения подпруги со спинным элементом (ремнем) установлено полукольцо для поводка или вожжей. 2. Упряжь по п.1, отличающаяся тем, что спинной элемент выполнен цельнокроеным и имеет форму трезубца, в зубцы которого жестко вшита подпруга, при этом спинной элемент выполнен многослойным и между его слоями размещена вставка из синтетического материала, а в местах скрепления зубцов с подпругой размещены полукольца для вожжей. 3. Упряжь по п.1 или 2, отличающаяся тем, что она дополнительно содержит карманы, для грузов, жестко ... 2180475 Устройство для поштучной подачи предметов, в частности семян сельскохозяйственных культур ... и достижения указанного выше технического результата, но при этом не исчерпывает всех возможностей осуществления изобретения, охарактеризованного совокупностью признаков, приведенной в формуле изобретения. Формула изобретения 1. Устройство для поштучной подачи предметов, в частности семян сельскохозяйственных культур, включающее в себя накопительный бункер для подаваемых предметов, бесконечную ленту для направленного выбрасывания предметов и приводное устройство, при этом накопительный бункер выполнен с возможностью вращения вокруг своей вертикальной оси под действием упомянутого приводного устройства, а вдоль, по меньшей мере, части периметра бункера выполнена щель, ширина ... 2403708 Устройство для полива сельхозрастений ... является устройство (журнал «Изобретатель и рационализатор», 12, 1973, стр.38 «Насос состоит из двух деталей»), содержащее вращающийся барабан, неподвижно закрепленную в центре трубку Пито, подводной и отводной трубопроводы. Недостаток - нет элементов в конструкции устройства для нагрева воды, подаваемой из реки для полива сельхозрастений. Цель изобретения - подогрев воды, подаваемой из реки в водонапорный резервуар.Поставленная цель достигается за счет того, что водяное колесо кинематически соединено с компрессором, теплогенератором, гидронасосом, состоящим из наружного барабана и внутреннего барабана, соединенных между собой перегородками; на ... |
Еще из этого раздела: 2165134 Корнеподрезающий рабочий орган машины для добычи лакричного сырья 2057432 Биологический состав кузнецова для подсочки деревьев, в том числе каучуконосов (варианты), и способ его приготовления 2050096 Мотокосилка 2196403 Почвообрабатывающий модуль 2494593 Способ повышения селена в чесноке горной зоны 2167648 Средство для защиты от укусов кровососущих насекомых (варианты) и способ его получения 2278488 Способ создания пастбищных экосистем весенне-летнего срока использования 2248352 Замещенные бензоилциклогександионы, гербицидное средство на их основе, исходное соединение 2062564 Способ оценки устойчивости растений к засухе северного и южного типа на ранних этапах онтогенеза 2076594 Установка для промышленного разведения дождевых червей |
Изобретения в сельском хозяйстве
Обработка почвы в сельском и лесном хозяйствах
Посадка, посев, удобрение
Уборка урожая, жатва
Обработка и хранение продуктов полеводства и садоводства
Садоводство, разведение овощей, цветов, риса, фруктов, винограда, лесное хозяйство
Новые виды растений или способы их выращивания
Производство молочных продуктов
Животноводство, разведение и содержание птицы, рыбы, насекомых, рыбоводство, рыболовство
Поимка, отлов или отпугивание животных
Консервирование туш животных, или растений или их частей
Биоцидная, репеллентная, аттрактантная или регулирующая рост растений активность химических соединений или препаратов
Хлебопекарные печи, машины и прочее оборудование для хлебопечения
Машины или оборудование для приготовления или обработки теста
Обработка муки или теста для выпечки, способы выпечки, мучные изделия
|
|
||
 3' вдоль нетраскрибируемой цепочки ДНК (т.е. цепочки, последовательность которой гомологична мРНК).
3' вдоль нетраскрибируемой цепочки ДНК (т.е. цепочки, последовательность которой гомологична мРНК).