Рекомбинантный стабильный окклюдированный вирус ядерного полиэдроза (варианты), инсектицидная композиция и способ борьбы с насекомыми на участкеПатент на изобретение №: 2156300 Автор: Патриция Джейн КЕЙЛЕЙ (GB), Лорна Мэри СТЮАРТ ДАЙТ (GB), Роберт Дэвид ПОССИ (GB), Мигель ФЕРБЕР ЛОПЕС (ES) Патентообладатель: Руссель-Юклаф (FR), Нэтчурал Энвайронмент Рисерч Каунсыл (GB) Дата публикации: 20 Сентября, 2000 Начало действия патента: 20 Марта, 1992 Адрес для переписки: 129010, Москва, ул. Большая Спасская 25, стр.3, ООО "Городисский и Партнеры", Лебедевой Н.Г. Изображения  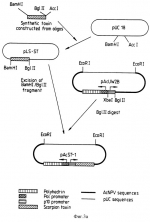 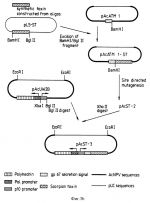 Настоящее изобретение относится к бакуловирусным инсектицидам. Предложены стабильные окллюдированные рекомбинантные вирусы ядерного полиэдроза, которые в клетках инфицированного ими насекомого экспрессируют токсин скорпиона или паука Segestria. Токсин выделяется из клеток насекомого. Такие рекомбинантные вирусы могут быть использованы для борьбы с насекомыми. Предложена инсектицидная композиция, содержащая инертный носитель и стабильный рекомбинантный окклюдированный вирус. Также предложен способ борьбы с насекомыми на участке, предусматривающий обеспечение на участке эффективного количества стабильного окклюдированного рекомбинантного вируса. 10 с.п. ф-лы, 19 ил., 4 табл. , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , ОПИСАНИЕ ИЗОБРЕТЕНИЯ К ПАТЕНТУНастоящее изобретение относится к бакуловирусным инсектицидам. Специфические к насекомым бакуловирусы представляют альтернативу применению химических агентов для борьбы с насекомыми в сельском хозяйстве. Они использовались для этих целей в течение многих лет. Бакуловирусы отличаются преимуществами селективности по отношению к мишеневым организмам и тем, что не загрязняют окружающую среду. Считают, что они безопасны для использования в программах борьбы с насекомыми в полевых условиях (WHO, 1973). Их основным недостатком является то, что они относительно медленно умерщвляют мишеневое насекомое, что мешает быстрому прекращению урона, наносимого в результате питания насекомых. Развитие методик генетических построений в области бакуловирусов дало возможность предположить, что эффективность таких агентов можно повысить за счет экспрессии в хозяина специфичных для насекомого токсинов, гормонов или энзимов, снижая тем самым время, необходимое для умерщвления насекомого, или, по крайней мере, предотвращая пожирание ими растений. До сих пор попытки модификации эффективности бакуловирусных инсектицидов имели ограниченный успех. Corbonell et al. (1988) экспрессировал специфический для насекомых токсин скорпиона из Buthus eupeus, используя Autographa californica вирус ядерного полиэдроза (AcMNPV), но не смог определить биологическую активность рекомбинантного токсина. Maeda (1989) включил ген диуретического гормона из Manduca sexta в Bombyx mori MNPV, и сокращает время, необходимое для умерщвления хозяина примерно на 20%. Hammock et al. (1990) экспрессировал эстеразу ювенильного гормона в Trichoplusia ni личинки, используя полиэдрин-негативный AcMNPV, и определил существенное снижение прироста веса при высоких дозах вируса. И, наконец, Merryweather et al. (1990) и Martens et al. (1990) включили ген дельта эндотоксина Bacillus thuringiensis в AcMNPV и получили аутентичный бактериальный токсин, который, хотя и был токсичен для личинки насекомого per se, не модифицировал эффективности вируса. Мы в настоящее время включили синтетический ген специфического к насекомому токсина под контролем бакуловирусного промотора в рекомбинантный AcMNPV. Мы обнаружили, что существует заметное повышение патогеничности к насекомому, если последовательность, кодирующую токсин, слить с последовательностью, кодирующей сигнальный пептид, который вызывает секрецию протеина. Изобретение относится к стабильному окклюдированному рекомбинантному вирусу ядерного полиэндроза, пригодному для использования в качестве инсектицида, причем вирус содержит один из следующих конструктов: - конструкт вектора pAcST3, представленный на фиг. 4в, - конструкт вектора pAcJL2ST, представленный на фиг. 9, - конструкт вектора pAcjHEST, представленный на фиг. 10, - конструкт вектора pAcSTST, представленный на фиг. 11, - конструкт вектора pAcCP2ST, представленный на фиг. 12, - конструкт вектора pAcOXYST, представленный на фиг. 13, - конструкт вектора pAcBPST, представленный на фиг. 14, - конструкт вектора pAcSefJT, представленный на фиг. 19. В настоящем изобретении предложен также способ борьбы с насекомыми на участке, который включает доставку на указанный участок количества рекомбинантных бакуловирусов по способу настоящего изобретения. Насекомое переваривает рекомбинантные бакуловирусы, вирусная ДНК мультиплицируется внутри насекомого, и токсин экспрессируется и выделяется, уничтожая насекомое. Гибель насекомого происходит быстрее, нежели при использовании рекомбинантных бакуловирусов, которые не снабжены последовательностью секреторного сигнала. Это обеспечивает важное преимущество, так как снижает количество растений, поврежденных насекомыми до их гибели. Настоящее изобретение можно применить к любым подходящим бакуловирусам семейства Baculoviridae, например, рода Baculovirus. Бакуловирусы могут быть вирусами ядерного полиэдроза (NPV), например, AcMNPV, Bombyx mori NPV, Heliothis zea NPV и Buzura suppressuria NPV. Геном рекомбинантного NPV по способу настоящего изобретения включает функциональный ген полиэдрина. Поэтому экспрессией проиэдринового протеина обычно управляет промотjр природного бакуловирусного полиэдрина. Так как рекомбинантные бакуловирусы являются функциональными в окружающей среде, эти вирусы должны быть жизнеспособными и должны обладать способностью создавать себе защитную оболочку. Геном рекомбинантных бакуловирусов включает также химерный ген, кодирующий инсектицидный токсин, или его функциональное производное, сигнальный пептид которого управляет секрецией структурного гена из клеток насекомого, инфицированного рекомбинантным бакуловирусом. Поэтому геном рекомбинантных бакуловирусов может состоять из дикого типа генома, в который включен химерный ген и элементы контролирующие его транскрипцию и трансляцию. Обычно на генетический механизм дикого типа генома не влияет добавление химерного гена с его контрольными последовательностями. Экспрессия протеинов, природно кодируемых дикого типа геномом, при этом не затрагивается. Таким образом, получают жизнеспособный, стабильный, инфекционный рекомбинантный бакуловирус. Токсин может быть нейротоксином, обычно нейротоксином, воздействующим на клеточные ионные каналы. Подходящим токсином является токсин, который выделяется клетками, которые нормально продуцируют токсин. Токсин обычно является полипептидным токсином ядовитого животного или микроорганизма. Подходящий токсин после очистки до гомогенности является токсичным для насекомого при введении. Предпочтительно, чтобы токсин действовал селективно против насекомых и не оказывал действия на млекопитающих. Рекомбинантные бакуловирусы могут эксперссировать и секретировать два или более токсина. Предпочтительным классом токсинов являются полипептиды, которые секретируются из клеток, в которых они синтезированы внутри животного в мешочек с ядом, из которого токсин вводится другому животному при укусе или ужалении. Примерами подходящих токсинов скорпиона являются следующие: 1. Токсины скорпиона, специфично действующие на насекомых. (тип Arthropoda. Класс Arachnida. Отряд Scorpionida) Buthus eupeus (Среднеазиатский скорпион) (см. последовательность 1 в конце текста). Последовательности I3A и I4A родственны I1A. Androctonus australis Hector (AaH IT): см. фиг. 1. Два других специфичных к насекомому токсина с родственными аминокислотными последовательностями были выделены из этого вида AaH IT1 и AaH IT2 (Loret, 1990) (см. последовательность 2 и 3). Было обнаружено, что этот токсин имеет две изоформы l, содержащие Ile в положениях 15 и 2 Val. (Lester, D., Lazarovici, P., Pelhate, M., Ziotkin, E. , Biochim. Biophys. Acta701, 370, 1982) (см. последовательность 4 и 5). 2. Токсин паука Segesdria florentina (Sef IT) может быть также использован (см. последовательность 6). Сигнальный пептид обеспечивает секретирование токсина из клеток, в которых он продуцируется. Сигнальная последовательность может быть последовательностью, способной обеспечивать секрецию протеина из клеток млекопитающих или насекомых, поэтому она может быть такой последовательностью млекопитающего, как секреторная сигнальная последовательность для интерлейкина 2 (IL-2), или такая последовательность насекомого, как секреторная сигнальная последовательность эстеразы ювенильного гормона, или протеин кутикулы Дрозофилы. Наиболее подходящей является бакуловирусная сигнальная последовательность, например, такая как секреторная сигнальная последовательность gp 67 протеина. Подходящие сигнальные последовательности поэтому могут включать (см. посл-ти 7-12). Ссылки (см. в конце текста). Сигнальный пептид может отщепляться от токсина, и токсин может быть секретирован. Поэтому сигнальный пептид можно сконденсировать с N-терминальным или C-терминальным аминокислотным остатком токсина либо непосредственно, либо за счет линкерной последовательности. Линкерная последовательность может содержать от 1 до 10 аминокислотных остатков, например, от 1 до 5 остатков. Рекомбинантные бакуловирусы настоящего изобретения можно получить за счет: (i) клонирования химерического гена, кодирующего чужой протеин, содержащий токсин, или его функциональное производное, и сигнальный пептид, в вектор переноса бакуловируса по рестрикционному сайту в прямом направлении от промотора, способного управлять экспрессией чужого протеина в клетки насекомого; и (ii) котрансфекции клеток, подверженных бакуловирусной инфекции с рекомбинантным вектором переноса со стадии (ii) и интактной ДНК дикого типа бакуловируса. Происходящая гомологическая рекомбинация приводит к получению рекомбинантного бакуловируса, содержащего химерический ген, операбельно связанный с промотором. Химерический ген обычно выделяют для использования на стадии клонирования (i). Рекомбинантные бакуловирусы могут также содержать экспрессируемый полиэдриновый ген. Этого можно достичь, используя на стадии (i) дикого типа бакуловирус, который включает такой ген. В другом варианте вектор переноса можно использовать на стадии (ii), который, кроме того, содержит последовательность кодирующего полиэдрин, операбельно связанную с промотором пролиэдрина. В этом случае на стадии (ii) можно использовать дикого типа бакуловирус без экспрессируемого гена полиэдрина. Предпочтительно, поэтому чтобы рекомбинантный бакуловирус содержал природный ген полиэдрина. Другими словами, последовательности, кодирующие полиэдрин и полиэдриновый промотор, являются природными для рекомбинантных бакуловирусов. Химерный ген содержит кодирующую последовательность для токсина. Эту кодирующую последовательность или полный химерический ген можно синтезировать. Олигонуклеотиды, которые в целом соответствуют целевым последовательностям, синтезируют и отжигают. Для этой цели предпочтительно адаптируют кодон, предпочтительный для конкретного бакуловируса, в который нужно включить кодирующую последовательность. Кодирующую последовательность, или полный структурный ген можно синтезировать с дополнительным 3"-терминальным кодоном для глицина, или эквивалентной последовательностью, которую после трансляции можно модифицировать до - NH2. Обеспечивается трансляционный стоп кодон. Ген токсина можно клонировать, используя известные специалистам методики. Если ген токсина синтезирован отдельно, его лигируют с соответствующей секреторной сигнальной кодирующей последовательностью. Секреторная сигнальная последовательность присутствует для того, чтобы обеспечить секрецию токсина из клеток насекомого, инфицированного рекомбинантным бакуловирусом настоящего изобретения. Предполагаемую сигнальную последовательность можно тестировать, конструируя рекомбинантные бакуловирусы, инфицируя их Spodoptera frugiperda клетками и исследуя надосадочную жидкость культур, инфицированных клетками S.frugiperda для секретированного токсина. В другом варианте гемолимфу можно выделить из насекомых инфицированных рекомбинантным бакуловирусом, ввести в чувствительные к ним насекомые и оценить токсичность. Токсин можно вводить как личинкам, так и взрослым особям. Для токсина, активного против двукрылых, тестовым насекомым может служить М. domestica. Другие подходящие насекомые можно выбрать в качестве тестовых видов для других токсинов при необходимости. Химерный ген содержит последовательности, контролирующие транскрипцию и трансляцию, включая промотор P10. Промотор управляет экспрессией протеина в клетках насекомого, инфицированного рекомбинантным бакуловирусом. Клетки, подверженные бакуловирусной инфекции, трансфектируют рекомбинантным вектором переноса, содержащим химерический ген, и инфицируют бакуловирусом дикого типа на стадии (ii), указанной ранее. Такие подверженные клетки обычно являются клетками клеточной линии насекомых, обычно клетками Spodoptera frugiperda. После того как происходит гомологическая рекомбинация, можно выделить рекомбинантный бакуловирус по способу настоящего изобретения. Рекомбинантный бакуловирус является стабильным. Другими словами, рекомбинантный бакуловирус достаточно стабилен для практических целей. Полиэдрин-негативный NPV не является достаточно стабильным для практических целей. Рекомбинантный бакуловирус является также инфекционным. Рекомбинантный бакуловирус может инфицировать любое подверженное ему насекомое. Чужой протеин, содержащий токсин экспрессируют в клетки насекомого и он секретируется из них. Токсин вызывает гибель насекомого. Поэтому рекомбинантный бакуловирус используют для борьбы с насекомыми-вредителями. Эффективное количество вносят на участок, например, на участок, где растут культурные виды или сорняки, которые служат хозяевами для вредителей-насекомых, и конкретные культурные растения, которые страдают от заражения насекомыми-вредителями. В другом варианте рекомбинантные бакуловирусы можно наносить на насекомых, например, содержащихся и выращиваемых в лаборатории насекомых, которых затем переносят на участок для инфицирования последующих поколений. Уровень дозы рекомбинантного вируса составляет от 108 до 1013 PIB на акр. Подходящий бакуловирус выбирают в зависимости от насекомого, подлежащего эрадикации. Рекомбинантный бакуловирус обычно наносят распылением, но его можно наносить в виде приманки или дуста. Рекомбинантный бакуловирус переваривается личинками насекомых, поэтому его следует вносить на соответствующий участок в то время, когда ожидается вылупление из яиц насекомого. Этот период длится обычно от двух до четырех недель. Вирус можно использовать отдельно или в сочетании с другими биологическими агентами контроля или химическими инсектицидами. В зависимости от выбранного бакуловируса такой способ борьбы можно использовать по отношению к насекомым следующих отрядов: Coleoptera Diptera Hymenoptera Neuroptera Trichoptera, и особенно, Lepidoptera. За счет выбора соответствующего бакуловируса таким способом можно контролировать множество насекомых вредителей важных культурных растений и лесов; причем насекомые включают следующие: Neodiprion sertifer Estigmene acrea Pectinophora gossypiella Plutella xylostella Arqyresthia conjugella Cydia molesta C. pomonella Pieris brassicae Manducca sexta Heliothis armigera, zea, virescens Mamestra brassicae Spodoptera littoralis, exigua, litura, exempta, mauritia Trichoplusia ni Diparopsis watersi Рекомбинантные бакуловирусы настоящего изобретения можно самим последовательно использовать в качестве инсектицидов обычным способом, в котором можно использовать немодифицированные нативные вирусы, например, внося эффективное количество на участок, где находятся насекомые подверженные инфицированию этим вирусом. Было обнаружено, что рекомбинантные бакуловирусы жизнеспособны или компетентны для таких целей благодаря тому, что содержат нативные ДНК последовательности для репродукции, летального инфицирования и экспрессии полипептида токсина. Рекомбинантные вирусы можно сконструировать таким образом, чтобы они экспрессировали и секретировали два или более из токсинов. Инсектицидные композиции, содержащие рекомбинантные бакуловирусы в инсектицидно эффективном количестве обычно содержат такие инертные носители, как глина, лактоза и протеинсодержащие материалы, например обезжиренный соевый порошок, которые способствуют нанесению, особенно такие протеинсодержащие носители, которые являются пищевыми аттрактантами, и которые могут повысить стабильность содержащихся в композиции бакуловирусов. Такие композиции желательно хранить при низких температурах, например, при 14 - 20oC для обеспечения сохранности вирусов. Композиции, предложенные в настоящем изобретении, могут быть измельченными композициями в том смысле, что они могут состоять из множества мелких частиц, в которых бакуловирусы сохраняются в глиняной и/или протеиновой матрице. Термин "сохраняются" использован здесь в том смысле, что такие частицы охватывают включенные в матрицы части бакуловирусов, экспонированных на поверхности частиц, а также включают полную инкапсуляцию бакуловирусов в матрицы, причем последнее предпочтительно, и является преимущественно для частиц настоящего изобретения. Размер частиц в композиции предпочтительно должен составлять не более 150 микрон, и следующей отличительной особенностью настоящего изобретения являются удовлетворительные композиции, с размерами "частиц от 5 до 100, предпочтительно, от 5 до 50 микрон, и особенно, от 5 до 25 микрон, которые достаточно легко получить. Материал матриц составляет от 65 до 99,9% по весу от полного веса бакуловируса и материалов матрицы и в предпочтительном варианте изобретения от 85 до 99 вес.%. Глина, которую можно использовать в рассматриваемой композиции, может быть любой удобно обрабатываемой глиной с мелкими порошкообразными частицами, например каолином, аттапульгитными и бентонитными глинами. Предпочтительными глинами являются оланча и аттапульгиты. Растительный протеин, который используют в качестве материала для матриц, также может быть любым из широкого круга источников протеина, которые имеют тонко измельченную форму. Эти материалы, предпочтительно, обезжиривают, или в другом варианте они практически не содержат жиров. Растительные источники протеина, которые можно указать в качестве иллюстрации, включают соевые бобы, семена хлопка, подсолнечника и экстракты различных дрожжей. Предпочтительными растительными протеинсодержащими материалами являются протеины сои и семян хлопка, предпочтительно из обезжиренного источника, более предпочтительно обезжиренные соевые бобы. Представительными животными протеинами являются снятое молоко, казеин и яичный альбумин. Протеинсодержащие материалы, получаемые из природных источников, могут содержать значительные количества непротеиновых материалов, и термины "протеин" и "протеинсодержащие материалы" используют здесь для обозначения материалов, которые содержат даже столь мало, как около, по крайней мере, 25 вес.% реального протеина. Весьма подходящие материалы, такие как предпочтительные растительные протеины, обычно содержат от 40 до 75% реального протеина. Хотя отдельные частицы и вся композиция могут содержать относительно низкие концентрации бакуловируса, желательно, с точки зрения минимальной практической пользы, чтобы композиция содержала активный эквивалент ЛД50 по крайней мере, 1,0 мкг/мл. Эффективность композиции может проявляться в дозах в интервале от 0,001 мкг/мл вплоть до 1,0 мкг/мл, и обычно в интервале от около 0,003 мкг/мл до 0,4 мкг/мл. Определение инсектицидной активности или эффективности, которые используют и на которые ссылаются, основаны на определении величин ЛД50, выраженной в мкг/мл питания, требующегося для обеспечения уровня доз для 50% роста личинок первой стадии при температуре 30oC. Этот метод подробно описан в J. Insect Pathology; 6,737-45 (1965) в отношении к оценке эффективности Trichopusia NPV. Измельченные композиции по способу настоящего изобретения демонстрируют необходимое количество инсектицидной эффективности, и обычно характеризуются повышенной сопротивляемостью к эффективности разложения за счет фотоинактивации и термоденатурации. Другие свойства композиций, особенно физические характеристики, меняются в зависимости от различных факторов, включая, в частности, материалы, используемые для матриц. Композиции, в которых матрицы состоят только из глины, обычно характеризуются хорошей смачиваемостью, но имеют тенденцию к меньшей слипаемости, и могут в некоторых случаях состоять из более мягких частиц. С другой стороны, композиции, в которых матрица состоит только из растительного протеина, могут быть в общем охарактеризованы хорошей прилипаемостью и твердостью, но обладают худшей смачиваемостью. Композиции, содержащие большие количества животного протеина, обладают хорошей прилипаемостью, но плохой смачиваемостью и жесткостью. Соответственно, предпочтительные материалы матриц выбирают из группы, состоящей из растительного протеина, глины и их смесей. Было также обнаружено, что композиции, в которых матрица состоит из гомогенной смеси растительного протеина и глины обладает преимуществами хорошей смачиваемости, слипаемости и жесткости, а также другими желательными характеристиками. Соответственно, наиболее предпочтительными композициями настоящего изобретения являются матрицы, состоящие из смеси растительных протеинов и глины. Весовое отношение растительного протеина к глине может быть в интервале от 0,1 до 10 вес. частей протеина на вес. часть глины и предпочтительно в интервале от 0,3 до 4 вес. частей растительного протеина на часть глины, более предпочтительно, от 0,5 до 3 вес. частей на часть глины. Бакуловирусы настоящего изобретения могут быть инкапсулированы в биоразлагаемом полимере, например, альгинате Culigel (торговая марка). В полимер может быть включен экран для ультрафиолетового света и/или кормовой стимулятор. Измельченный солнцезащитный агент, например, углерод, красители на основе углерода, алюминийоксид, двуокись титана, глина, мука или флуоресцентные материалы могут присутствовать в полимере. Нижеследующие примеры иллюстрируют изобретение. В соответствии с прилагаемыми рисунками: На фиг. 1 представлена AcMNPV gp67 сигнальная последовательность и AaHIT кодирующая последовательность, gp67 сигнальный пептид выделен двумя потенциальными кодонами, инициирующими трансляцию жирным шрифтом. На фиг. 2 представлена последовательность синтетических олигонуклеотидов, использованных при конструировании AaHIT гена. Нуклеотидные последовательности представлены в 5"-3"-направлении (a1, b3, c5 и d7) ив направлении 3"-5" (a2, b4, c6 и d8). Сайты рестрикционных энзимов, критические для сборки гена токсина, представлены над последовательностью. Кодон инициирования трансляции подчеркнут, а кодон остановки трансляции (TAA) отмечен звездочкой над вторым нуклеотидом. "Stuffer" участок между BamH1 и Acc1 сайтами включен для облегчения двойного перевара этих двух сайтов, необходимых при конструировании. Олигонуклеотиды включены последовательно в сайт поликлонирования pUC18. d7/d8 были включены между Acc1 и Xba1 сайтами и pUC18 (Сайт Acc18 сконструирован для разрушения при вставке), образуя pSTD. c5/c6 включают между BamH1 и Xba1 сайтами pSTD для образования pSTDC. Это переваривают BamH1 и Acc1 для высвобождения "Stuffer" участка, и a1/a2 клонируют в эти сайты для создания pSTDCA. Это переваривают Acc1 и вставляют b3/b4 для создания pLS-ST. На фиг. 3а изображено конструирование pAcST-1. Синтетические олигонуклеотиды, кодирующие AaHIT, были собраны в pUC18, как показано на фиг. 2, до получения pLS-ST. Затем полный AaHIT, кодирующий участок, удаляют из pLS-ST, и вставляют в Bgl II сайт pAcUW2B, вектор переноса бакуловируса, содержащий копию p10 промотора, расположенную в прямом направлении полного гена полиэдрина AcMNPV. Полученный вектор переноса pAcST-1 содержит полную AaHIT кодирующую последовательность без сигнального пептида. На фиг. 3b представлено конструирование pAcST-3. Синтетический AaHIT удаляют из pLS-ST и вставляют по BamH I сайту pAcATM1 для получения pAcATV-1-ST. Мутагенез под действием сайта осуществляют по связи между gp67 сигналом секреции и AaHIT кодирующим участком до получения благоприятного протеазного сайта расщепления в модифицированной плазмиде pAcST-2. Модифицированный gp67-AaНIТ кодирующий участок удаляют из pAcST-2, и вставляют в pAcUW2B для получения pAcST-3. На фиг. 4A представлена нуклеотидная последовательность AcMNPV p10 промотора (1-240) в ассоциации с gp67 сигнальным пептидом, кодирующим участок (246-359) и AaHIT кодирующий участок (360-569). Сайт инициирования транскрипции из p10 промотора (положение 167-170; TAAG) указан жирным шрифтом. Связь между сигнальным пептидом и токсином показана до мутагенеза. Нуклеотиды, которые были удалены, показаны ниже. На фиг. 4b представлена нуклеотидная последовательность AcMNPV p10 промотор гена в соответствии с gp67 сигнальным пептидным кодирующим участком и AaHIT кодирующим участком. Над последовательностью представлена рестрикционная карта плазмиды вектора переноса (pAcST3), которую использовали для получения рекомбинантного вируса. На фиг. 5 представлена экспрессия протеинов в клетки, инфицированные вирусом S. frigiperda (Sf). Эти клетки были инфицированы AcST-3 или вирусом дикого типа, и импульсно-мечены 35S цистеином в течение 1 часа в различные моменты после инфицирования, как описано в методике. Протеины выделяли на 10-30% полиакриламидном градиентном геле перед экспонированием пленки рентгеновским лучам. Указано положение AaHIT и протеина полиэдрина. На фиг. 6 представлены результаты иммуноблотнинг анализа ВЖХ (HPLC) фракции AcH полного токсина и AcST-3 инфицированных клеток среды. Фракции 25-28 из ВЖХ были выделены на 10-30% полакриламидном градиентном геле и были подвергнуты блоттингу, как указано в тексте. BSA (присутствующие в повторно суспендированном буфере) и AaHIT были указаны. На фиг. 7 представлены полный неочищенный AaH яд или среда из Sf клеток, инфицированных AcST-3, которые были фракционированы и использованы в ВЖХ (см. пример 1). Анализ выбранных фракций осуществляли, вводя 4 мкл в M.domestica и наблюдая паралич или гибель. На фиг. 8 представлено поверхностное поражение листьев личинкой, инфицированной AcST-3 и AcMNPV (дикого типа). На фиг. 9 представлена нуклеотидная последовательность AcMNPV p10 генного промотора в ассоциации с участком, кодирующим сигнальный пептид интерлейкина 2, и AaHIT кодирующий участок. Над этой последовательностью изображена рестрикционная карта плазмиды вектора переноса (pAcI12ST), использованной для получения рекомбинантного вируса. На фиг. 10 представлена нуклеотидная последовательность AcMNPV p10 генного промотора в ассоциации с участком, кодирующим сигнальный пептид эстеразы ювенильного гормона и AaHIT кодирующий участок. Над этой последовательностью представлена рестрикционная карта плазмиды вектора переноса (pAcJHEST), использованной для получения рекомбинантного вируса. На фиг. 15 представлен синтез протеинов в инфицированных вирусом клетках Sf, как представлено на фиг. 5 (а) ACI12ST; (b) AcJHEST; (c) AcBPST. На фиг. 16 представлена AcMNPV gp67 сигнальная последовательность и SefIT кодирующая последовательность. gp67 сигнальный пептид выделен двумя потенциальными кодонами инициирования трансляции жирным шрифтом. На фиг. 17 представлена последовательность синтетических олигонуклеотидов, использованных при конструировании SefIT гена. Нуклеотиды представлены в направлении 5"-3" (OS3711) и в направлении 3"-5" (OS3712). Сайты рестрикционных энзимов, использованных для субклонирования пар олигонуклеотидов в плазмиды, представлены ниже последовательности. Кодон инициирования трансляции подчеркнут, а кодон остановки трансляции (TAG) указан звездочкой над вторым олигонуклеотидом. После отжига олигонуклеотиды включают в pDH7, между Sph I и BamH I сайтами. pDH7 является производным pEMBL19, который имеет сигнал секреции AcNPV gp67 между уникальным Bgl II сайтом, расположенным вместо Hind III сайта полилинкера, и Sph I. На фиг. 18 представлена нуклеотидная последовательность AcMNPV h10 промотора (1-240) в ассоциации с gp67 участком, кодирующим сигнальный пептид (246-359). Остальная последовательность является участком, кодирующим SefIT. Последовательность представлена после мутагенеза связи между gp67 участком и SefIT участком. На фиг. 19 представлена нуклеотидная последовательность AcMNPV p10 генного промотора в ассоциации с участком, кодирующим сигнальный пептид gp67, и SefIT кодирующим участком. Над последовательностью представлена рестрикционная карта плазмиды вектора переноса (pAcSefIT), использованной для получения рекомбинантного вируса. Пример 1. Материалы и способы (A) Вирусы и клетки AcMNPV C6 (Possee, 1986) и все рекомбинантные вирусы культивируют в Spodoptera frugiperda (IPLB-Sf-21) клетках (Vaughn et al., 1977) при 28oC в TC100 с добавлением 5% или 10% сыворотки плода теленка (FCS) в присутствии пенициллина и стрептомицина. Инфицирующую способность вируса оценивали, используя анализ бляшкообразования по способу Brown and Faulkner (1977). Очистка вируса: Полиэдру очищают из инфицированных клеток или насекомых по способу Harrap et al. (1977), но с добавлением 0,1% (вес/объем) SDS ко всем растворам. SDS удаляют, промывая полиэдру водой в конце процесса очистки. Количество полиэдры оценивают, используя способ сухого счета Wigley (1976). (B) Конструирование синтетических AaHIT и SefIT кодирующих последовательностей Протеиновую последовательность AaHIT превращают в нуклеотидную последовательность, используя смещение предпочтительного кодона AcMNPV полиэдрина (Hooft van Iddenkinge et al, 1983) и p10 (Kuzio et al, 1984) генов. Ожидаемую ДНК последовательность анализируют на VAX компьютере, используя программы молекулярной биологии Стадена для идентификации сайтов рестрикционного энзима. Их используют для конструирования четырех комплементарных пар синтетических олигонуклеотидов, которые получают на автоматическом синтезаторе Applied Biosystem. Последовательность конструируют таким образом, чтобы четыре нуклеотида перед ATG кодоном соответствовали нуклеотидам AcMNPV гена полиэдрина, сохраняя, таким образом, целостность микроокружения инициирования трансляции; кодон окончания трансляции включают по 3" концу кодирующего участка. Соответствующие сайты рестрикционного энзима включают для облегчения включения синтетических олигонуклеотидов в pUC18 и их последующего вырезания в виде целого AaHIT кодирующего участка BamHI и Bgl II для последующих манипуляций. Олигонуклеотиды собирают в полную AaHIT кодирующую последовательность с помощью pUC18 (Yanisch-Perron et al., 1985), используя стандартные протоколы (Sambrook et al., 1989). Дальнейшие подробности этой процедуры описаны ранее в отношении фиг. 2, 3а и 3b. AaHIT кодирующий участок в окончательной конструкции плазмиды, pLS-ST, секвенируют, используя терминаторы дидеоксинуклеотидных цепей (Sanger et al., 1977) для подтверждения их аутентичности. Аналогичным образом протеиновую последовательность Sef IT превращают в информационную нуклеотидную последовательность. Синтезируют два комплементарных олигонуклеотида, OS3711 и OS3712 (фиг. 17). После отжига эти олигонуклеотиды включают в плазмиду pDH7, между сайтами Sph I и BamH I. pDH7 является производным pEMBL19 (Dente et al., 1983), которое содержит сигнал секреции AcMNPV gp67 между уникальным Bgl II сайтом, расположенным вместо Hind III сайта полилинкера, и Sph I. Sef IT кодирующий участок в конструкции плазмиды, pDH7-Sef IT, секвенируют описанным ранее способом для подтверждения его аутентичности. (C) Конструирование векторов переноса бакуловируса (i) pAcST-1 (сравнение): Для получения практичного вируса-инсектицида необходимо, чтобы AaHIT кодирующая последовательность была включена в бакуловирус, который сохраняет функциональный ген полиэдрина. Вектор переноса pAcUW2B (Weyer et al. , 1990) удовлетворяет этому критерию. Он основан на AcMNPV EcoR I-I фрагменте, который включает ген полиэдрина. Он имеет полную копию гена полиэдрина, копию p10 промотора и сайт клонирования Bgl II с SV 40 последовательностью окончания транскрипции, включенной в положение на 90 нуклеотидов в обратном направлении от полиэдринового ATG кодона и в противоположной ориентации. AaHIT кодирующий участок иссекают из pLS-ST, используя BamH I и Bgl II, очищают после фракционирования в геле, желирующемся при низкой температуре, и включают в Bgl II сайт вектора переноса рАcUW2B (Weyer et al., 1990) до получения pAcST-I (фиг. 3a). (ii) pAcST-2: Для получения вирусов, которые потенциально могут секретировать AaHIT кодирующий участок, включают в pAcATMI, вектор переноса с копией gp67 сигнальную пептидную последовательность (Whitford et al., 1989) после промотора полиэдрина. Участок, кодирующий токсин, иссекают из pLS-ST с помощью BamH I и Bgl II и включают в BamH I сайт в pAcATMI, получая вектор переноса pAcST-2 (фиг. 3b). pAcATMI является производным pAcCL29 (Livingstone and Jones, 1989), вектором переноса на базе промотора полиэдрина, содержащем М13 межродовой участок, который облегчает получение однонитевой ДНК за счет использования М13 хелперного фага (Livingstone and Jones, 1989). Связь между gp67 сигнальной последовательностью и AaHIT модифицируют, используя М13 олигонуклеотидный направленный мутагенез (Kunkel, 1985), используя GP-67-SC. Tox, изображенный на фиг. 4A до получения соответствующей ДНК последовательности, кодирующей сайт узнавания подходящей протеазы: на этой стадии также удаляют исходный кодон инициирования трансляции ATG от AaHIT. Изменения, осуществленные в gp67 соединении, представлены на фиг. 4А наряду с последовательностью p10 промотора. (iii) pAcST-3 (изобретение): gp67 сигнальную последовательность и AaHIT удаляют из pAcST-2, переваривая Xho II, и помещая в Bgl II сайт в pAcUW2B до получения pAcST-3 (фиг. 3b). (iv) pAcl12ST (изобретение): AaHIT синтетический кодирующий участок включают как BamH I / Bgl II переваренный ДНК фрагмент в BamH I сайт с полилинкерным участком BLUESCRIPT М13 phagemid (Stratagene). Ориентацию кодирующего участка подтверждают рестрикционным энзимным картрированием. Последовательность, кодирующую сигнальный пептид IL-2 (Taniguchi et al., 1963), включают между EcoR V/Pst I сайтами в обратном направлении от кодона инициирования трансляции последовательности токсина. Соединение между последовательностями кодирующими сигнальный пептид и токсин, изменяют, используя олигонуклеотидный-направленный мутагенез (Kunkel, 1985) для получения оптимального сайта распознавания протеазы для отщепления секреторного пептида. Последовательность подтверждают, используя методы терминации цепи. ДНК фрагмент, содержащий участки, кодирующие сигнальный пептид и токсин, иссекают с помощью Xho II и включают по Bgl II сайту вектора переноса бакуловируса pAcUW21 до получения pAcI12ST. рAcUW21 является производной pAcUW2B и имеет укороченные вирусные последовательности, фланкирующие ген полиэдрина и p10 генный промотор; она также содержит соответствующую регуляторную последовательность для обеспечения продуцирования однонитевой ДНК в бактериях, после суперинфицирования за счет M13 KO7. pAcUW21 конструируют, иссекая нуклеотидный фрагмент ДНК 5408 из pAcUW2B и включая его в pEMBL19, которая была переварена Hind III и EcoR I, причем концы ДНК восстановлены ДНК полимеразой и обработаны до гилирования фосфатазой кишечника теленка. На фиг. 9 представлены последовательность участка, кодирующего I1-2 сигнальный пептид, в ассоциации с AaHIT кодирующей последовательностью. (v) pAcJHEST (изобретение): Синтетический ДНК кодирующий JHE сигнальный пептид конструируют, включая две пары перекрывающихся олигонуклеотидов в полилинкер модифицированного варианта pEMBL19 (сайт Hind III был изменен до Bgl II) между Bgl II и EcoR I сайтами. Затем эту плазмиду переваривают BamH I (внутри 3" участка JHE последовательности), участок, кодирующий токсин, включают как BamHI/Bgl II-переваренный ДНК фрагмент. Связь между JHE сигнальным пептидом и участками, кодирующими токсин меняют, используя олигонуклеотид-направленный мутагенез, как было указано ранее. Затем эти последовательности выделяют Bgl II перваром и включают в pAcUW21 по Bgl II сайту до получения pAcJHEST. На фиг. 10 представлен участок, кодирующий сигнальный пептид JHE в ассоциации с AaHIT кодирующей последовательностью. (vi) pAcSTST (изобретение): Для получения копии AaHIT гена с сигнальной пептидной последовательностью для природного AaHIT получают методом полимеразной цепной реакции Bougis Р.Е., et al., 1989) по 5" концу и Bgl II с обоих концов. 5" праймер (см. в конце описания последовательность 13). 3" праймер: TATATAAGATCT TTA GTT AAT AAT AGT AGT GTC Затем переваренный Bgl II фрагмент вставляют в pAcUW21 по уникальному Bgl II сайту. Этот вектор обозначают pAcSTST. На фиг. 11 представлены AaHIT сигнальный пептид и AaHIT кодирующий участок. (vii) pAcCP2ST (изобретение): Этот вектор получают, как и для pAcSTST, за исключением того, что добавляют сигнальную пептидную последовательность для CP2 из Drosophila melongaster (Syder, M. et al., 1982) вместо природного AaHIT сигнального пептида. 5" праймер имеет вид (см. посл-ть 14 в конце описания). 3" праймер имеет вид: TATATAAGATCT TTA GTT AAT AAT AGT AGT GCT. На фиг. 12 представлены последовательность сигнального пептида и AaHIT кодирующего участка. (viii) pAcOXYST (изобретение): Этот вектор получают как и для pAcSTST, за исключением сигнальной пептидной последовательности для окситоцина из крыс (IvelI, R. and Tichter, D., 1984) добавляют вместо природного AaHIT сигнального пептида. 5" праймер имеет вид (последовательность 15) 3" праймер имеет вид: TATATAAGATCT TTA GTT AAT AAT AGT AGT GTC. Последовательность окситоцинового сигнального пептида и AaHIT кодирующего участка представлены на фиг. 13. (ix) pAcUW21 BP (изобретение): Промотор основного протеинового гена иссекают из pAcMPI (Hill-Perkins et al., 1990) с помощью ASp 718 и BamH I и вставляют между теми же сайтами в pDH7 для получения pDH7BP. Метод полимеразной цепной реакции используют затем для получения копии промотора основного протеинового гена, а также добавляют Xba I сайт по 5" концу промоторного фрагмента. Затем этот фрагмент вставляют вместо p10 генного промотора в pAcUW21 и обозначают pAcUW21BP. ДНК последовательность, содержащую gp67 сигнальный пептид и участок, кодирующий токсин, иссекают из pAcST-3, используя Xho II и включают по Bgl II сайту pAcUW21BP для получения pAcBPST. Последовательности основного протеинового и p10 генного промотора в ассоциации с gp67 и AaHIT кодирующими участками изображены на фиг. 14. (х) pAcSefIT (изобретение): gp67 кодирующий участок помещают в рамку с SefIT кодирующим участком (в pDH7-SefIT), используя сайт направленного мутагенеза, как было указано ранее. Изменение подтверждают секвенированием. Полный кодирующий участок (gp67 и SefIT) иссекают из pDH7-SefIT с помощью Bgl II и BamH I и включают по Bgl II сайту pAcUW21 до получения pAcSefIT. Последовательность SefIT кодирующего участка в ассоциации с gp67 сигнальным пептидным участком и p10 генным промотором представлена на фиг. 18 и 19. (D) Получение рекомбинантных вирусов (i) AcST-1 (сравнение): Spodoptera frugiperda клетки были совместно трансфектированы с pAcST-1 и полиэдрин гегативной AcRP-6-SC (Kitts et al., 1990) инфекционной вирусной ДНК, используя кальций фосфатную процедуру (Smith et al. , 1983). Рекомбинантные вирусы выбирают как полиэдрин-позитивный фенотип в стандартном анализе блашкообразования. (ii) AcST-3 (изобретение): pAcST-3 совместно трансфектируют с AcRP6-SC, используя метод липофектина (Felgner et al., 1987, Groebe et al., 1990), и вирусы скринируют как для предыдущей котрансфекции. (iii) AcI12ST, AcJHEST, AcBPST, AcSTST, AcCP2ST, AcOXYST, AcSefIT (все изобретение): все описаны как для AcST-3. Рекомбинантные вирусы очищают пятью последовательными анализами бляшкообразования и определяют их аутентичность Саузерн гибридизацией EcoR I переварами ДНК из инфицированных клеток, используя [32p] меченые олигонуклеотиды, специфичные для AaHIT гена в качестве зонда. (F) Иммуноблоттинг протеинов инфицированных клеток или фракций ВЖХ (высокоэффективная жидкостная хроматография) Протеиновые экстракты немеченых, инфицированных вирусом клеток (см. E ранее или G далее) выделяют в 10-30% денатурирующие полиактиламидные гели, и переносят на нитроцеллюлозные фильтры, используя аппаратуру электроблоттинга Bio-Rad в течение двух часов при 100 В в 25 мМ-трис, 5,ЗМ-глицине и 20% метаноле в качестве буфера переноса. Фильтр инкубируют в PBS, содержащем 0,05% Твин-20 (PBST) с 3-5% жирности порошком молока в качестве блокирующего агента. Все это осторожно встряхивают в течение 1 часа при комнатной температуре, затем инкубируют еще в течение 2 часов с сывороткой морской свинки против синтетического AaHIT пептида, разбавленного 1/100 в PBST (титр более чем 1: 10000 в косвенном ELISA против гомологичного пептида). Затем фильтр промывают несколько раз в PBST для удаления несвязанных антител. Связанные антитела детектируют, используя антиморская свинка lgG-щелочной фосфатазный конъюгат. (G) ВЖХ AaH полного токсина и инфицированных клеточных надосадочных жидкостей, и in vivo анализ в Musca domestica: тест на токсичность при инъекции Полный неочищенный AaH яд, полученный от Sigma в виде замороженного высушенного образца (1 мг), ресуспендируют в 20 мкл ДМСО, затем разбавляют до 100 мкм в H2O. Выпавший в осадок материал удаляют центрифугированием при 15000 g в течение 5 минут. Очистку надосадочной жидкости (20 мкл) ведут с помощью ВЖХ, используя LCD систему с детектором Spectroflow 280 нм. Для разделения используют Spherisord, C8, с диаметром частиц 5 мкм в 25 см колонке. В качестве растворителя A используют 0,15 M аммонийформиат. pH 2,7 и растворитель В -ацетонитрил. Элюирование ведут с линейным градиентом от 15 до 40% в A за 40 минут при скорости потока 1 мл/мин, и собирают 40 фракций по 1 мл. Фракции сушат вымораживанием для удаления ацетонитрила, и формиата аммония, затем ресуспендируют в 200 мкл 0,1 мг/мл BSA в воде. По 0,5 мкл каждой фракции вводят 10 взрослым M.domestica (самцам и самкам), используя иглы шприца 25 gauge. Анализ надосадочной жидкости от инфицированных клеток проводят, используя надосадочную жидкость с 2 чашек с клетками инфицированных S.frugiperda в качестве исходного материала (то есть, 2 мл). Это концентрируют в 4 раза, используя систему фильтрации Centricon (ЗК) за счет центрифугирования при 500 g в течение 3 часов на центрифуге Sorvall RC5. 100 мкл концентрированного материала загружают в колонку ВЖХ, и обрабатывают так же, как и полный яд. После обработки образца на ВЖХ, фракции сушат вымораживанием и ресуспендируют в 100 мкл 0,1 мг/мл BSA. По 4 мкл каждой фракции используют для введения M.domestica. (Н) Получение вирусов в насекомых T.ni выдерживают в лаборатории на полусинтетической диете (Hunter et al. , 1984). Все анализы проводят, используя полиэдру, которую один раз пропустили через насекомых. Для начального проведения, третьей стадии T.ni личинкам давали корм, который содержал неочищенный экстракт инфицированных клеток, после чего личинки переводят на свежую диету. Сразу после гибели насекомых их собирают и обрабатывают. Полиэдру очищают и используют для последующих анализов. (1) Биологическая активность гемолимфы, выделенной из инфицированных клеток Биологические активности токсина оценивают, вводя по 4 мкл серийных разбавлений конкретного образца в dorso lateral участок брюшка M.domestica, между склерой. Фракции, которые содержат токсин, полученный из ВЖХ для aAH полного яда, используют для создания стандартов для оценки количества токсина, выделенного из гемолимфы. Фракции сушат вымораживанием и снова суспендируют до концентрации 90 нг/100 мкл воды. Это снова разбавляют в PBS для облегчения введения 4 мкл, содержащих 18, 9, 4,5, 2,25 или 1,25 нг токсина насекомым (доза 18 нг оказалась избыточной для ЛД50 для токсина). (J) Определение ЛД50 Во второй стадии личинок T. ni (примерно 0,6-0,7 мг) отдельно кормят пятью сериальными разбавленными известного титра вируса на небольших кусочках искусственного корма в микротитровальных пластинах. Максимальная доза вируса в идеале должна приводить к более 90% гибели, а минимальная доза должна приводить к 10% гибели. He-инженерные штаммы AcMNPV удовлетворяющие этим условиям, составляют 120, 60, 30, 15 и 7,5 PIB на личинку. Те личинки, которые употребили дозу за 24 часа, и те, которые не употребили, были исключены из анализа. Остальных перенесли в индивидуальные контейнеры с искусственным кормом и выдерживали в течение 24 часов в темном инкубаторе. Личинок ежедневно осматривали и кадавров удаляли, помещали на предметное стекло, и причину гибели определяли под микроскопом. Полученные результаты анализировали, используя probit анализ для определения величин ЛД50 (Finney, 1971). (K) Определение ST50 Для получения новорожденных (neonate) личинок, которых используют в таких исследованиях, взрослых особей помещают в ячейки, содержащие фильтровальные бумажки для откладки яиц. Фильтры с яичками поверхностно стерилизуют и сохраняют в пластиковых контейнерах. После вылупления новорожденным не дают пищи в течение 3-6 часов, перед тем, как дают капельную пищу с различными суспензиями вирусов. Суспензии подкрашивают 5% синим пищевым красителем для визуализации корма. Маленькие капли этой суспензии помещают в чашки петри концентрическими кольцами. Личинки помещают в центр таких концентрических окружностей, после чего перемещаются через капли, отбирая небольшой объем жидкости до того, как они вползают на крышку чашки. Личинки, которые оказываются поврежденными в процессе манипуляций, остаются в центре чашки, что позволяет синхронно кормить и отбирать здоровые особи. В предварительных работах было показано, что в этом типе экспериментов личинка T. ni потребляет 0,0087 ( 0,0023) мкл (Hughes and Wood, 1981. Hughes et al., 1986) и в этих ST50 анализах суспензии вирусов содержали 1 0,0023) мкл (Hughes and Wood, 1981. Hughes et al., 1986) и в этих ST50 анализах суспензии вирусов содержали 1 106 полиэдры/мл. После кормежки личинки содержались в отдельных контейнерах с искусственным кормом. Спустя 24 часа личинки осматривают и все погибшие личинки удаляют, так как их гибель связана с манипуляциями; спустя еще 48 часов личинки еще раз осматривают, и затем интервалы между осмотрами сокращают. Все погибшие личинки удаляют и причину гибели определяют по внешнему виду и при изучении под микроскопом. Оценку ST50 проводят по программе Vistat (Boyce Thompson Institute, Ithaca. New York). (L) Снижение повреждений, наносимых при кормежке насекомыми инфицированными AcST-3 Личинок третьей стадии (T.ni) кормят отдельно в течение ночи кусочками искусственного корма инокулированного 104 телами включения полиэдры (PIB). Столь высокая доза приводит к 100% гибели. После кормежки личинки разделяют на группы по 5, на листах капусты, которая выращивалась в контролируемых условиях. Перед кормежкой листья погружают на ночь в воду для обеспечения насыщения. После двух дней кормежки повреждение листьев оценивают и фотографируют. 2. Результаты (A) Синтез протеинов рекомбинантными вирусами AaHIT имеет протеиновую последовательность, содержащую 70 аминокислот (включая первый метионин) с предсказанным молекулярным весом 8000. Анализ экстрактов из клеток S.frugiperda клеток инфицированных AcST-1 (AaHIT без секреторного сигнала) или AcST-3 (AaHIT с секреторным сигналом) в денатурирующем 10-30% полиакриламидном геле, в котором не определяется дополнительных протеинов ожидаемого размера, либо окрашиванием Comassie Blue или серебром. Затем инфицированные вирусом клетки метят [35S] цистеином в различные времена после инфицирования (фиг. 5). Результаты показывают дополнительный протеин, который мигрирует как раз между ожидаемым положением вирусного p10 протеина (p10 протеин без цистеиновых остатков). Синтез предполагаемых AaHIT максимален между 18 и 36 часами после инфицирования. Идентичность протеина подтверждают с использованием Вестернблоттинг анализа AcST-1 или AcST-3 инфицированных клеточных экстрактов с антисывороткой, доведенной до синтетической AaHIT. (B) ВЖХ AaH полного яда и синтетического AaHIT На хроматограммах ВЖХ видны пики с оптической плотностью на 280 нм вокруг фракций 27/28, которые присутствуют в полном яде, и среде клеток, инфицированных AcST-3, но отсутствуют в клетках инфицированных дикого типа вирусами. Вестернблоттинг анализ фракций между 25 и 28 указывает на наличие AaHIT во фракциях 27 и 28 неочищенного яда, и в 26 27 надосадочной жидкости клеток, инфицированных AcST-3 (фиг. 6). Инъекция M. domestica небольшими объемами каждой из этих фракций подтверждает, что для этих фракций наблюдается паралитическая и летальная активности. Иммобилизация за счет токсина наблюдается через час после инъекций, а гибель становится очевидной через 24 часа после инъекций (фиг. 7). (C) Биологическая активность гемолимфы, выделенной из инфицированных личинок Секрецию AaHIT из AcST-3 инфицированных клеток in vivo оценивают, выделяя гемолимфу из инфицированных вирусом T.ni личинок и вводя ее в M.domestica. Личинки 4 стадии T.ni инфицируют 105 PIB AcST-3. Спустя 72 часа после инфицирования производят надрез у концевой ложноножки и отбирают гемолимфу (20 мкл). Для ограничения меланизации, пипетки, используемые для сбора гемолимфы всех образцов, погружают в раствор фенилтиомочевины. По 4 мкл этой гемолимфы используют для инъекций тестовым личинкам M.domestica и оценивают токсичность. Реакцию сравнивают со стандартными кривыми, полученными с использованием AaHIT полного яда очищенного на ВЖХ, а полученные результаты представлены в таблице 1. Реакция M.domestica указывает концетрации токсина в личинках T. ni около 1,5-2,0 нг/мкл гемолимфы спустя 72 часа после инфицирования. (D) In vivo активности рекомбинантных вирусов In vivo активности рекомбинантных вирусов оценивают, определяя величины ЛД50. Инфицируют личинки T.ni второй стадии. Повторным особям вводят дозы 7, 15, 30, 60 или 120 PIB. После probit анализа ЛД50 величины для немодифицированного AcMNPV составили 44 (95% предел доверительности 35 и 55), AcST-1 38 (95% пределы доверительности 31 и 46) и для AcST-3 (изобретение) - 31 (95% пределы доверительности 27 и 37). ЛД50 значения для AcMNPV и AcST-3 заметно отличается для 106 полиэдры/мл. После кормежки личинки содержались в отдельных контейнерах с искусственным кормом. Спустя 24 часа личинки осматривают и все погибшие личинки удаляют, так как их гибель связана с манипуляциями; спустя еще 48 часов личинки еще раз осматривают, и затем интервалы между осмотрами сокращают. Все погибшие личинки удаляют и причину гибели определяют по внешнему виду и при изучении под микроскопом. Оценку ST50 проводят по программе Vistat (Boyce Thompson Institute, Ithaca. New York). (L) Снижение повреждений, наносимых при кормежке насекомыми инфицированными AcST-3 Личинок третьей стадии (T.ni) кормят отдельно в течение ночи кусочками искусственного корма инокулированного 104 телами включения полиэдры (PIB). Столь высокая доза приводит к 100% гибели. После кормежки личинки разделяют на группы по 5, на листах капусты, которая выращивалась в контролируемых условиях. Перед кормежкой листья погружают на ночь в воду для обеспечения насыщения. После двух дней кормежки повреждение листьев оценивают и фотографируют. 2. Результаты (A) Синтез протеинов рекомбинантными вирусами AaHIT имеет протеиновую последовательность, содержащую 70 аминокислот (включая первый метионин) с предсказанным молекулярным весом 8000. Анализ экстрактов из клеток S.frugiperda клеток инфицированных AcST-1 (AaHIT без секреторного сигнала) или AcST-3 (AaHIT с секреторным сигналом) в денатурирующем 10-30% полиакриламидном геле, в котором не определяется дополнительных протеинов ожидаемого размера, либо окрашиванием Comassie Blue или серебром. Затем инфицированные вирусом клетки метят [35S] цистеином в различные времена после инфицирования (фиг. 5). Результаты показывают дополнительный протеин, который мигрирует как раз между ожидаемым положением вирусного p10 протеина (p10 протеин без цистеиновых остатков). Синтез предполагаемых AaHIT максимален между 18 и 36 часами после инфицирования. Идентичность протеина подтверждают с использованием Вестернблоттинг анализа AcST-1 или AcST-3 инфицированных клеточных экстрактов с антисывороткой, доведенной до синтетической AaHIT. (B) ВЖХ AaH полного яда и синтетического AaHIT На хроматограммах ВЖХ видны пики с оптической плотностью на 280 нм вокруг фракций 27/28, которые присутствуют в полном яде, и среде клеток, инфицированных AcST-3, но отсутствуют в клетках инфицированных дикого типа вирусами. Вестернблоттинг анализ фракций между 25 и 28 указывает на наличие AaHIT во фракциях 27 и 28 неочищенного яда, и в 26 27 надосадочной жидкости клеток, инфицированных AcST-3 (фиг. 6). Инъекция M. domestica небольшими объемами каждой из этих фракций подтверждает, что для этих фракций наблюдается паралитическая и летальная активности. Иммобилизация за счет токсина наблюдается через час после инъекций, а гибель становится очевидной через 24 часа после инъекций (фиг. 7). (C) Биологическая активность гемолимфы, выделенной из инфицированных личинок Секрецию AaHIT из AcST-3 инфицированных клеток in vivo оценивают, выделяя гемолимфу из инфицированных вирусом T.ni личинок и вводя ее в M.domestica. Личинки 4 стадии T.ni инфицируют 105 PIB AcST-3. Спустя 72 часа после инфицирования производят надрез у концевой ложноножки и отбирают гемолимфу (20 мкл). Для ограничения меланизации, пипетки, используемые для сбора гемолимфы всех образцов, погружают в раствор фенилтиомочевины. По 4 мкл этой гемолимфы используют для инъекций тестовым личинкам M.domestica и оценивают токсичность. Реакцию сравнивают со стандартными кривыми, полученными с использованием AaHIT полного яда очищенного на ВЖХ, а полученные результаты представлены в таблице 1. Реакция M.domestica указывает концетрации токсина в личинках T. ni около 1,5-2,0 нг/мкл гемолимфы спустя 72 часа после инфицирования. (D) In vivo активности рекомбинантных вирусов In vivo активности рекомбинантных вирусов оценивают, определяя величины ЛД50. Инфицируют личинки T.ni второй стадии. Повторным особям вводят дозы 7, 15, 30, 60 или 120 PIB. После probit анализа ЛД50 величины для немодифицированного AcMNPV составили 44 (95% предел доверительности 35 и 55), AcST-1 38 (95% пределы доверительности 31 и 46) и для AcST-3 (изобретение) - 31 (95% пределы доверительности 27 и 37). ЛД50 значения для AcMNPV и AcST-3 заметно отличается для  = 0,05. Различие в ЛД50 относительно невелико, причем возрастание составляет 1,4191 с 95% пределами доверительности 1,026216-1,9846. ST50 оценивают после инфицирования новорожденных. Однако демонстрируют весьма заметное снижение во временах, необходимых для умерщвления личинок. ST50 для AcMNPV составило 113,1 (95% доверительные пределы 112-115), для AcST-1 - 109,2 (95% пределы доверительности 107-110) и для AcST-3 (изобретение) составило 85,8 часов (95% пределы доверительности 85-87 часов). Результат для AcST-3 заметно отличается от значений для немодифицированного вируса AcST-1, который содержит токсин кодирующую последовательность без секреторного сигнала = 0,0001). Это означает, что AcST-3 вирус убивает T.ni личинки, по крайней мере, на 25% быстрее, нежели AcMNPV. Полученные результаты суммированы в таблице 2. (E) Патология инфицирования Исследовали также патологию после инфицирования AcMNPV, AcST-1, или AcST-3. Личинки, инфицированные AcST-3, отличаются от личинок, инфицированных AcST-1 или AcMNPV. Инфицирование AcST-1 или AcMNPV приводит к массированному вызванному вирусом повреждению, приводящему к разжижению личинок, сопровождающемуся кремоватой окраской. При инфицировании AcST-3 личинки сохраняют свою структурную целостность и остаются зеленого цвета. (F) Уменьшение повреждений наносимых питанием личинок, инфицированных AcST-3 Эффективность AcST-3 рекомбинантного вируса по снижению повреждений, наносимых культурным растениям T.ni личинками, представлена на фиг.4. (G) Синтез протеинов рекомбинантными вирусами с альтернативными сигнальными пептидами Экспрессию гена токсина, включенного в каждый из рекомбинантных вирусов, тестируют, инфицируя Spodoptera frugiperda клетки и вводя радиометки в отдельные культуральные чашки с одинаковыми временными интервалами в 1 час. По лученные результаты представлены на фиг. 15a, b. AcJHEST продуцируют некоторое количество токсина спустя 12 часов после инфицирования, с максимумом синтеза между 18-36 часа ми после инфицирования; после чего скорость продуцирования протеина снижается. Сравнимые результаты наблюдают для АсI12ST-инфицированных клеток, хотя выход токсина из клеток, инфицированных этим вирусом, может быть несколько ниже. (H) Синтез протеинов рекомбинантными вирусами с гибридным генным промотором На фиг. 15c представлены также кинетики продуцирования токсина скорпиона в AcBPST-инфицированных клетках. Очевидно, что вирус инициирует продуцирование токсина в значительных количествах спустя 12 часов после инфицирования. Выход значительно выше, нежели из любых других клеток, инфицированных рекомбинантными вирусами, содержащими ген токсина под контролем одного p10 генного промотора. Продуцирование токсина в AcBPST-инфицированных клетках продолжается вплоть до 48 часов после инфицирования. (I) Биологическая активность токсина, секретированного в среде от инфицированных вирусом клеток В таблице 3 представлены биологические активности AaHIT, секретированного в культуральную среду, поддерживающую рост инфицированных вирусом Sf клеток. Среду исследуют, используя M.domestica, как описано в (C). Очевидно, что AcST-1 не выделяет активный токсин в среду. (J) In vivo активность рекомбинантных вирусов с альтернативными сигналами секреции или гибридным генным промотором Величины ЛД50 для каждого рекомбинантного вируса оценивают, как описано ранее (D). Найдено, что они аналогичны. St50 оценивают после инфицирования новорожденных. ST50 для AcST-3 составляет 71,4 часа (95% пределы доверительности 70,6-72,2), для AcJHEST составляет 77,7 часа (95% пределы доверительности 76,7-78,7), для AcI12ST составляет 87,2 часа (95% пределы доверительности 86,1-86,3), для AcBPST составляет 73,1 часа (95% пределы доверительности 71,8-74,4). В таблице 4 приведены ЛД50 и ST50 для рекомбинантного бакуловируса, содержащего AaHIT. E. coli XZ-блю, содержащие pAcATM-1, E.coli HB101, содержащие pAcUW2B и E. coli SURETM, содержащие pAcST-3, были депонированы в Национальной Коллекции Промышленных и Морских Бактерий, Абердин, Великобритания, 22 марта 1991 г. под регистрационными номерами NCIMB 40393, NCIMB 40394 и NCIMB 40395, соответственно. = 0,05. Различие в ЛД50 относительно невелико, причем возрастание составляет 1,4191 с 95% пределами доверительности 1,026216-1,9846. ST50 оценивают после инфицирования новорожденных. Однако демонстрируют весьма заметное снижение во временах, необходимых для умерщвления личинок. ST50 для AcMNPV составило 113,1 (95% доверительные пределы 112-115), для AcST-1 - 109,2 (95% пределы доверительности 107-110) и для AcST-3 (изобретение) составило 85,8 часов (95% пределы доверительности 85-87 часов). Результат для AcST-3 заметно отличается от значений для немодифицированного вируса AcST-1, который содержит токсин кодирующую последовательность без секреторного сигнала = 0,0001). Это означает, что AcST-3 вирус убивает T.ni личинки, по крайней мере, на 25% быстрее, нежели AcMNPV. Полученные результаты суммированы в таблице 2. (E) Патология инфицирования Исследовали также патологию после инфицирования AcMNPV, AcST-1, или AcST-3. Личинки, инфицированные AcST-3, отличаются от личинок, инфицированных AcST-1 или AcMNPV. Инфицирование AcST-1 или AcMNPV приводит к массированному вызванному вирусом повреждению, приводящему к разжижению личинок, сопровождающемуся кремоватой окраской. При инфицировании AcST-3 личинки сохраняют свою структурную целостность и остаются зеленого цвета. (F) Уменьшение повреждений наносимых питанием личинок, инфицированных AcST-3 Эффективность AcST-3 рекомбинантного вируса по снижению повреждений, наносимых культурным растениям T.ni личинками, представлена на фиг.4. (G) Синтез протеинов рекомбинантными вирусами с альтернативными сигнальными пептидами Экспрессию гена токсина, включенного в каждый из рекомбинантных вирусов, тестируют, инфицируя Spodoptera frugiperda клетки и вводя радиометки в отдельные культуральные чашки с одинаковыми временными интервалами в 1 час. По лученные результаты представлены на фиг. 15a, b. AcJHEST продуцируют некоторое количество токсина спустя 12 часов после инфицирования, с максимумом синтеза между 18-36 часа ми после инфицирования; после чего скорость продуцирования протеина снижается. Сравнимые результаты наблюдают для АсI12ST-инфицированных клеток, хотя выход токсина из клеток, инфицированных этим вирусом, может быть несколько ниже. (H) Синтез протеинов рекомбинантными вирусами с гибридным генным промотором На фиг. 15c представлены также кинетики продуцирования токсина скорпиона в AcBPST-инфицированных клетках. Очевидно, что вирус инициирует продуцирование токсина в значительных количествах спустя 12 часов после инфицирования. Выход значительно выше, нежели из любых других клеток, инфицированных рекомбинантными вирусами, содержащими ген токсина под контролем одного p10 генного промотора. Продуцирование токсина в AcBPST-инфицированных клетках продолжается вплоть до 48 часов после инфицирования. (I) Биологическая активность токсина, секретированного в среде от инфицированных вирусом клеток В таблице 3 представлены биологические активности AaHIT, секретированного в культуральную среду, поддерживающую рост инфицированных вирусом Sf клеток. Среду исследуют, используя M.domestica, как описано в (C). Очевидно, что AcST-1 не выделяет активный токсин в среду. (J) In vivo активность рекомбинантных вирусов с альтернативными сигналами секреции или гибридным генным промотором Величины ЛД50 для каждого рекомбинантного вируса оценивают, как описано ранее (D). Найдено, что они аналогичны. St50 оценивают после инфицирования новорожденных. ST50 для AcST-3 составляет 71,4 часа (95% пределы доверительности 70,6-72,2), для AcJHEST составляет 77,7 часа (95% пределы доверительности 76,7-78,7), для AcI12ST составляет 87,2 часа (95% пределы доверительности 86,1-86,3), для AcBPST составляет 73,1 часа (95% пределы доверительности 71,8-74,4). В таблице 4 приведены ЛД50 и ST50 для рекомбинантного бакуловируса, содержащего AaHIT. E. coli XZ-блю, содержащие pAcATM-1, E.coli HB101, содержащие pAcUW2B и E. coli SURETM, содержащие pAcST-3, были депонированы в Национальной Коллекции Промышленных и Морских Бактерий, Абердин, Великобритания, 22 марта 1991 г. под регистрационными номерами NCIMB 40393, NCIMB 40394 и NCIMB 40395, соответственно.
ФОРМУЛА ИЗОБРЕТЕНИЯ1. Стабильный окклюдированный рекомбинантный вирус ядерного полиэдроза для использования в качестве инсектицида, причем вирус содержит фрагмент ДНК, состоящий из следующих последовательно расположенных элементов: гена токсина скорпиона, последовательности ДНК, кодирующей сигнальный пептид gp67, промотора р10, гена полиэдрина с собственным промотором, и содержащий 2 сайта рестрикции XbaI, 1 сайт рестрикции BglII, 1 сайт рестрикции HindIII и 1 сайт рестрикции KpnI. 2. Стабильный окклюдированный рекомбинантный вирус ядерного полиэдроза для использования в качестве инсектицида, причем вирус содержит фрагмент ДНК, состоящий из следующих последовательно расположенных элементов: гена токсина скорпиона, последовательности ДНК, кодирующей сигнальный пептид IL-2, промотора р10, гена полиэдрина с собственным промотором, и содержащий 2 сайта рестрикции XbaI. 3. Стабильный окклюдированный рекомбинантный вирус ядерного полиэдроза для использования в качестве инсектицида, причем вирус содержит фрагмент ДНК, состоящий из следующих последовательно расположенных элементов: гена токсина скорпиона, последовательности ДНК, кодирующей сигнальную последовательность эстеразы ювенильного гормона, промотора р10, гена полиэдрина с собственным промотором, и содержащий 2 сайта рестрикции XbaI. 4. Стабильный окклюдированный рекомбинантный вирус ядерного полиэдроза для использования в качестве инсектицида, причем вирус содержит фрагмент ДНК, состоящий из следующих последовательно расположенных элементов: гена токсина скорпиона, последовательности ДНК, кодирующей сигнальную последовательность AaHIT, промотора р10, гена полиэдрина с собственным промотором, и содержащий 2 сайта рестрикции XbaI, 1 сайт рестрикции BamHI, 1 сайт рестрикции HindIII, 1 сайт рестрикции KpnI. 5. Стабильный окклюдированный рекомбинантный вирус ядерного полиэдроза для использования в качестве инсектицида, причем вирус содержит фрагмент ДНК, состоящий из следующих последовательно расположенных элементов: гена токсина скорпиона, последовательности ДНК, кодирующей сигнальную последовательность СР2; промотора р10, гена полиэдрина с собственным промотором, и содержащий 2 сайта рестрикции XbaI, 1 сайт рестрикции BamHI, 1 сайт рестрикции HindIII и 1 сайт рестрикции KpnI. 6. Стабильный окклюдированный рекомбинантный вирус ядерного полиэдроза для использования в качестве инсектицида, причем вирус содержит фрагмент ДНК, состоящий из следующих последовательно расположенных элементов: гена токсина скорпиона, последовательности ДНК, кодирующей сигнальную последовательность окситоцина, промотора р10, гена полиэдрина с собственным промотором, и содержащий 2 сайта рестрикции XbaI, 1 сайт рестрикции BamHI, 1 сайт рестрикции HindIII и 1 сайт рестрикции KpnI. 7. Стабильный окклюдированный рекомбинантный вирус ядерного полиэдроза для использования в качестве инсектицида, причем вирус содержит фрагмент ДНК, состоящий из следующих последовательно расположенных элементов: гена токсина скорпиона, последовательности ДНК, кодирующей сигнальную последовательность gp 67, промотора р10, промотора гена основного белка AcMNPV, гена полиэдрина с собственным промотором, и содержащий сайт рестрикции XbaI. 8. Стабильный окклюдированный рекомбинантный вирус ядерного полиэдроза для использования в качестве инсектицида, причем вирус содержит фрагмент ДНК, состоящий из следующих последовательно расположенных элементов: гена токсина паука Segestria, последовательности ДНК, кодирующей сигнальную последовательность gp 67, промотора р10, гена полиэдрина с собственным промотором, и содержащий 1 сайт рестрикции BgIII, 1 сайт рестрикции XbaI, 1 сайт рестрикции BamHI, 1 сайт рестрикции HindIII, 1 сайт рестрикции KpnI. 9. Инсектицидная композиция, содержащая инертный носитель и активный компонент, отличающаяся тем, что в качестве активного компонента содержит стабильный рекомбинантный окклюдированный вирус по пп.1 - 8 в эффективном количестве. 10. Способ борьбы с насекомыми на участке, предусматривающий обеспечение на участке эффективного количества инсектицидно-активного вирусного препарата, отличающийся тем, что в качестве вируса используют стабильный окклюдирующий рекомбинантный вирус по пп.1 - 8.Популярные патенты: 2106081 Животноводческая ферма с применением помещений круглой формы и способ содержания в ней, например, крупного рогатого скота ... тем, что групповая автопоилка установлена на центральной опоре, выполнена в виде кольцевой чаши с ребрами жесткости и снабжена механизмом в виде червячного редуктора с тросовыми барабанами для установки ее по высоте. 5. Ферма по п.1, отличающаяся тем, что кормораздатчик установлен на центральной опоре, выполнен в виде корытообразной емкости с цепочно-планчатым транспортером и совмещен с электропастухом, выполненным в виде массивной рамки оголенного электропровода низкого напряжения, повешенного на тросик и закрепленного на вращающихся барабанах. 6. Секции животноводческой фермы для молодняка крупного рогатого скота с применением помещений круглой формы, включающая ... 2027341 Бункер для сыпучих материалов ... в виде пневмотрубы и снабжен струйным аппаратом, расположенным в каждом вышеназванном отсеке. Цилиндрический корпус в отсеках, примыкающих своей плоскостью к стенке корпуса, выполнен перфорированным и снаружи закрыт кожухами с образованием кольцевых полостей вокруг перфорации. Нижняя перегородка образует с днищем коническую полость. Кроме того, бункер снабжен циклоном, встроенным в его верхнюю часть, причем входной патрубок циклона выполнен в виде спиральной трубы, установленной в верхней части пневмотрубы. Струйный аппарат выполнен с возможностью перекрытия всасывающих патрубков. Бункер снабжен приводом для элементов перекрытия зазоров конических перегородок и патрубков струйных ... 2484613 Способ создания почвенно-растительного покрова при рекультивации нарушенных земель ... наносят механизированным или ручным способом (при небольших размерах участка) 5 см насыпной слой из промышленных песчаных отходов, содержащих питательные и известковые элементы, которые экранируют растения от загрязненного грунта, нейтрализуют его высокую кислотность, повышают его питательный статус и иммобилизуют тяжелые металлы, что снижает фитотоксичность грунта. Насыпной слой может быть больше и увеличен до 10 см. Его дополнительно удобряют минеральными удобрениями - N70 P70K70. Затем вручную или механизированным способом (например, используя сеялки для посева семян или разбрасыватели твердых удобрений) рассыпают по поверхности слоя отходов подготовленный вспученный ... 2231250 Устройство для промышленного выращивания земляники и других растений ... распирающими трубами. Верхняя распирающая труба имеет множество отверстий по длине растительного поля (не показаны) для подачи питательного раствора, а также вентили и/или штуцеры 6, служащие, кроме того, и для фиксации растительного поля на опорной раме (не показана). На трубе выполняются и другие штуцеры (не показаны) для подачи воздуха, пара и газовых смесей.Нижняя распирающая труба также имеет множество отверстий и вентили и/или штуцеры (не показаны) для отсасывания питательного раствора в резервуары регенерации (не показаны).На нижний край растительного поля навешивается поддон 7 со штуцером 8 для отсасывания питательного раствора, не захваченного нижней распирающей трубой. ... 2161402 Способ акселерационного содержания и разведения кроликов ... создать новый по качеству продукт, так как мясо таких крольчат при значительной массе имеет необыкновенно тонкий вкус и аромат, является молочным и отличается от привычного мяса кролика. Расселение самцов по персональным ячейкам по возможности по одиночке осуществляют преимущественно на 110-й день от рождения. Реализацию кроликов, достигших веса в 4-5 килограмма, на расплод, мех и мясо осуществляют с 122-го по 150-й день от рождения с учетом сезона, пола кролика и степени вызревания его меха. Срок вызревания меха кролика крайне непостоянен и зависит от ряда обстоятельств - календарной даты рождения кролика, количества крольчат на подсосе у крольчихи, порядкового номера окрола, ... |
Еще из этого раздела: 2120753 Способ получения пестицидного водного суспензионного концентрата и пестицидный водный суспензионный концентрат 2395497 Способ стимулирования роста подсолнечника регулятором роста 2402211 Способ получения трансгенных кроликов, продуцирующих белки в молочную железу 2250602 Широкозахватный колесный дождеватель 2215407 Способ создания исходного материала для селекции растений 2050096 Мотокосилка 2423042 Электронно-оптический способ регулирования технологии производства агропродукции 2295848 Способ дезинсекции и дезинфекции материалов зернового происхождения и устройство для его осуществления 2078495 Устройство для транспортирования кормов в хранилищах башенного типа 2152151 Гербицидная водорастворимая гранулированная композиция |
Изобретения в сельском хозяйстве
Обработка почвы в сельском и лесном хозяйствах
Посадка, посев, удобрение
Уборка урожая, жатва
Обработка и хранение продуктов полеводства и садоводства
Садоводство, разведение овощей, цветов, риса, фруктов, винограда, лесное хозяйство
Новые виды растений или способы их выращивания
Производство молочных продуктов
Животноводство, разведение и содержание птицы, рыбы, насекомых, рыбоводство, рыболовство
Поимка, отлов или отпугивание животных
Консервирование туш животных, или растений или их частей
Биоцидная, репеллентная, аттрактантная или регулирующая рост растений активность химических соединений или препаратов
Хлебопекарные печи, машины и прочее оборудование для хлебопечения
Машины или оборудование для приготовления или обработки теста
Обработка муки или теста для выпечки, способы выпечки, мучные изделия
|
|
||