Способ оценки влияния ингибитора деацетилирования белков на индукцию ростового морфогенеза растенийПатент на изобретение №: 2404585 Автор: Иванова Эвилина Алексеевна (RU), Вафина Гюльнар Хамидовна (RU) Патентообладатель: Учреждение Российский академии наук Институт биологии Уфимского научного центра РАН (ИБ УНЦ РАН) (RU) Дата публикации: 27 Ноября, 2010 Начало действия патента: 21 Апреля, 2009 Адрес для переписки: 450054, г.Уфа, пр. Октября, 69, Учреждение Российской академии наук Институт биологии Уфимского научного центра РАН, А.И. Мелентьеву Изображения  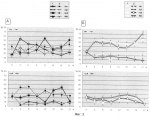 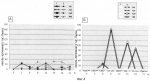 Изобретение относится к физиологии и биохимии растений. Первоначально проращивают семена пшеницы в 0,004 мМ растворе бутирата натрия, в определенные интервалы времени от начала замачивания 0 ч, 3 ч, 6 ч, 9 ч, 12 ч, 15 ч, 18 ч, 21 ч проводят отделение зародышей от эндосперма с последующей консервацией зародышей в забуференном 80-90% глицерине при минус 25°С, выделяют клеточные ядра, проводят экстракцию ядерных фракций возрастающими концентрациями 0,14 М, 0,35 М, 2 М хлористого натрия и 6 М гуанидин гидрохлорида с 0,1% Изобретение относится к физиологии и биохимии растений и может быть применено к анализу механизмов ростового морфогенеза растений, а именно при исследовании процессов, происходящих на уровне супраструктур интерфазного клеточного ядра, которые иногда называют «молекулярными морфогенезами», физико-химическая природа которых мало исследована, что необходимо для получения дополнительной информации в разработках и построении компьютерных моделей организации генных и эпигенных сетей управления. Известен способ оценки влияния гиббереловой кислоты и кинетина на белки клеточных ядер [1], в котором был описан способ оценки влияния физиологически активных веществ на белки клеточных ядер проростков растений. Недостаток этого метода заключается в том, что с его помощью невозможно оценить влияние физиологически активных веществ на индукцию ростового морфогенеза растений. Вышеуказанный способ оценки влияния гиббереловой кислоты и кинетина на белки клеточных ядер [1] был принят за основу, в котором первоначально осуществляют обработку семян в процессе набухания физиологически активными веществами, инкубацией в среде с 2-14С-ацетатом и Na2H32PO4, с последующей изоляцией клеточных ядер по методам Ро, Чипчейза и Кюля [см. 1], получением ядерных экстрактов растворами солей: 0,14 М, 2 М хлористого натрия, определением турбидиметрически содержание белка в пробах, фракционный состав белков определяли с помощью электрофореза. Недостаток этого метода заключается в том, что в анализ берется только один временной период роста растения, а именно 2 суток, что не позволяет оценить морфогенетические изменения в растении в процессе индукции ростовых процессов, происходящие под влиянием физиологически активных веществ, клеточные ядра выделяются методами Ро, Чипчейза и Кюля [см. 1], что ведет к низкому выходу нативных ядер, фракционирование клеточных ядер ограничивается получением только 2-х фракций, содержание белка в которых определяется турбидиметрически, т.е. методом с низким порогом чувствительности (до 3-5 мкг белка в 0,1 мл раствора). Цель изобретения - предлагается использование Арг-Х протеолиза, как одного из молекулярно-генетических механизмов крупномасштабной пространственной организации хроматина в условиях гиперацетилированного состояния протеома клеточных ядер при индукции ростового морфогенеза растений. Указанная цель достигается тем, что в способе оценки влияния ингибитора деацетилирования белков на индукцию ростового морфогенеза растений первоначально проращивают семена пшеницы в 0,004 мМ растворе бутирата натрия, в определенные интервалы времени от начала замачивания 0 ч, 3 ч, 6 ч, 9 ч, 12 ч, 15 ч, 18 ч, 21 ч проводят отделение зародышей от эндосперма с последующей консервацией зародышей в забуференном 80-90% глицерине при минус 25°С, выделяют клеточные ядра, проводят экстракцию ядерных фракций возрастающими концентрациями 0,14 М, 0,35 М, 2 М хлористого натрия и 6 М гуанидин гидрохлорида с 0,1% Изобретение иллюстрируется следующим примером. Пример. Опыты проводили на элитных семенах пшеницы (Thticum aestivum L.) сортов Мироновская 808 (озимая) и Мироновская яровая (любезно присланные нам из коллекции Мироновского научно-исследовательского института селекции и семеноводства пшеницы им. В.Н. Ремесло). Проращивание зародышей осуществляли в темноте при 22±1°С. Соответственно, в одном варианте опыта (контроль) использовали дистиллированную воду, в другом варианте - 0,004 мМ NaВ(бутират натрия), растворенный в дистиллированной воде. Бутират натрия использовали для ингибирования in vivo процессов деацетилирования гистонов (возможно и других негистоновых белков). То есть для поддержания удлиненной (пролонгированной) стадии повышенного уровня ацетилирования гистонов (возможно и других ядерных белков), необходимого для стимуляции транскрипции, NaB был синтезирован на базе Института органической химии УНЦ РАН. Элементный анализ бутирата натрия находился в пределах: С - 43,5; Н - 6,3; Na - 21. Оптимальную концентрацию NaB подбирали по увеличению всхожести семян пшеницы Мироновской яровой через 21 ч после индукции ростовых процессов (таблица). В определённые интервалы времени - 0 ч (воздушно-сухое семя) и от начала замачивания семян: 3 ч, 6 ч, 9 ч, 12 ч, 15 ч, 18 ч, 21 ч проводили отделение зародышей от эндосперма. Клеточные ядра выделяли по методу [2]. Надмолекулярные структуры: нуклеоплазму (Нп), хроматин непрочно- (Хр-I) и прочно- (Хр-II) связанный с ядерным матриксом (ЯМ), а также ЯМ выделяли из очищенных клеточных ядер соответственно при повышении ионной силы раствора: 0,14М NaCl, 0,35М NaCl, 2М NaCl в 0,01М трис-HCl буфере, рН 6.8. ЯМ извлекали 6М гуанидингидрохлоридом (Gu·HCI) с 0,004%-ным Анализ таблицы показал, что максимальная всхожесть семян на 21 ч прорастания соответствует концентрации бутирата натрия 0,004 мМ. На фиг.1 представлена всхожесть семян пшениц Мироновская яровая (А) и Мироновская озимая (Б), контроли (1), обработанные бутиратом натрия (2). I - не проклюнулись, II - проклюнулись: общий размер проростка 0,3-0,5 см, III - активный рост главного корня (более 0,1 см). На оси ординат показан процент всхожести семян. На фиг.2 представлена динамика внутриядерного протеома G1 фазы клеточных ядер зрелых зародышей Мироновской яровой (А) и Мироновской 808 (озимой) (Б) пшениц в нормальных условиях (1) и в присутствии ингибитора деацетилирования белков (2). На оси абсцисс показано время прорастания зародышей пшеницы. На оси ординат - масса 1 зародыша, мг. На фиг.3 представлен выход белковых компонентов во фракциях клеточных ядер зрелых зародышей Мироновской яровой (А) и Мироновской 808 (озимой) (Б) пшениц в нормальных условиях (1) и в присутствии ингибитора деацетилирования белков (2). Использованы следующие обозначения: Нп-нуклеоплазма, Хр-I - хроматин непрочносвязанный с ядерным матриксом, Хр-II - прочносвязанный с ядерным матриксом, ЯМ - ядерный матрикс. На оси абсцисс показан возраст зародышей пшеницы, по оси ординат - процент выхода белковых компонентов. На фиг.4 представлена Арг-Х протеолитическая активность во фракциях клеточных ядер зрелых зародышей Мироновской яровой (А) и Мироновской 808 (озимой) (Б) пшениц в нормальных условиях (1) и в присутствии ингибитора деацетилирования белков (2). Использованы следующие обозначения: Нп - нуклеоплазма, Хр-I - хроматин непрочносвязанный с ядерным матриксом, Хр-II - прочносвязанный с ядерным матриксом, ЯМ - ядерный матрикс. На оси абсцисс указан возраст зародышей пшеницы, по оси ординат - единицы протеолитической активности нмоль аргинина·с-1·мг·белка. Анализ фиг.4 показал, что протеазочувствительность супраструктур клеточного ядра в присутствии ингибитора деацетилирования ядерных белков резко влияет на экспонированность определенных участков ядерного матрикса (9 ч), нуклеоплазмы (15 ч) при сохраненности Арг-Х протеолиза на уровне ядерного матрикса в контрольном варианте опыта (18 ч). 18-ч фаза клеточного цикла у пшеницы интересна тем, что в этот временной период наблюдается репликация и переход к синтезу ДНК, т.е. клетка осуществляет процессы, необходимые для их сохранения в митозе. Данное изобретение рекомендуется для молекулярно-генетического анализа механизмов ростового морфогенеза растений, а именно при исследовании процессов, происходящих на уровне супраструктур интерфазного клеточного ядра.  Влияние NaB на всхожесть семян пшеницы сорта Мироновская яровая Концентрация бутирата Na (мМ) Всхожесть семян на 21 ч, % 080 0,004 920,01 88 0,01576 0,5 701 72 252 4 32 Влияние NaB на всхожесть семян пшеницы сорта Мироновская яровая Концентрация бутирата Na (мМ) Всхожесть семян на 21 ч, % 080 0,004 920,01 88 0,01576 0,5 701 72 252 4 32
Формула изобретенияСпособ оценки влияния ингибитора деацетилирования белков на индукцию ростового морфогенеза растений, включающий проращивание семян пшеницы в 0,004 мМ растворе бутирата натрия, отделение зародышей от эндосперма в определенные интервалы времени от начала замачивания: 0 ч, 3 ч, 6 ч, 9 ч, 12 ч, 15 ч, 18 ч, 21 ч, консервацию зародышей в забуференном 80-90% глицерине при минус 25°С, с последующим выделением клеточных ядер, экстракцией ядерных фракций возрастающими концентрациями 0,14 М, 0,35 М, 2 М хлористого натрия и 6 М гуанидин гидрохлорида с 0,1% Популярные патенты: 2277321 Колосоподъемник для косилочных систем уборочных машин ... со стеблеподъемником. Между первым и вторым концами на несущей линейке закреплен кронштейн с удерживающими средствами для опирания на косилочный палец. Кронштейн образует с удерживающими средствами проход для обеспечения возможности приближения несущей линейки к косилочному пальцу. Несущая линейка изготовлена из полосового материала и в зоне между вторым концом и кронштейном имеет большую жесткость, чем в зоне между первым концом и кронштейном. Повышенная жесткость обеспечивается за счет выполнения обращенного к стеблеподъемнику участка выпуклым с дугообразным сечением. Колосоподъемник обеспечивает ориентацию в сторону почвы конца несущей линейки в зоне закрепления со ... 2015654 Теплица для подземной выработки ... рамы 2. Каждая секция рамы 2 выполнена в виде многоярусной гидропонной установки, имеющей два вертикальных ригеля 3, жестко связанных балкой 4, имеющей с двух сторон консольные участки 5. Верхние секции многосекционной поперечной рамы 2 имеют только вертикальные ригели 6 и жестко закреплены в верхней части подземной выработки 1. Крайние боковые секции многосекционных поперечных рам 2 жестко закреплены в стенах выработки 1 посредством консолей 5 балок 4. Секции соседних многосекционных рам 2 жестко соединены между собой перекрытиями-полами 7. Вертикальные ригели 6, 3 многоярусных гидропонных установок имеют кронштейны 8 для контейнеров 9 с растениями, которые соединяют ригели ... 2201065 Приемная часть осевого сепаратора ... разрезе по линии V-V фиг.1 в конце приемной части. Сведения, подтверждающие возможность осуществления изобретения На фиг. 1 показана часть зерноуборочного комбайна 1, который содержит двухбарабанный молотильный аппарат 2 с тангенциальным направлением перерабатываемой массы, подающий барабан 3 и установленный вслед за подающим барабаном 3 осевой сепаратор 4, оснащенный приемной частью 4а в соответствии с изобретением. При этом вместо двухбарабанного молотильного аппарата 2 в комбайне может быть установлен любой однобарабанный или многобарабанный молотильный аппарат с тангенциальным направлением массы. Осевой сепаратор 4 установлен на комбайне с помощью известных средств ... 2437864 Способ микробиологической переработки птичьего помета ... агар) и на содержание аммонийного азота (Петербургский А.В. Практикум по агрономической химии. М.: Изд-во Сельхозлитературы, - 1963. - 592 с.) Результаты исследований представлены в таблице 2. Таблица 2 Зависимость биоконверсии помета кур от времени обработки культурой Pseudomonas sp.114 Время от начала засева, сутки ПОКАЗАТЕЛИ Общее микробное число в курином помете, кл/мл Аммонийный азот, мг/л 0106 250 5 107 г 19510 109 16515 1010 110 201010 115 30 1010 122По результатам испытаний отмечено, что наибольшее количество микробных клеток птичьего помета достигнуто к 15 суткам после внесения микробной культуры Pseudomonas sp.114.К 20 и 30 суткам количество ... 2195644 Монитор для определения качества зерна ... от 1 до 4%. Далее пространственно разделенные потоки с различными длинами волн поочередно фокусируются на приемник 52. Указанный приемник относится к типу, одновременно измеряющему выходной сигнал в широком интервале по длинам волн. В предпочтительном варианте осуществления приемник 52 представляет собой ПЗС- матрицу, ячейки (пикселы) которой индивидуально измеряют интенсивность излучения на каждой из соответствующих длин волн. Другими словами, каждая ячейка ПЗС-матрицы настраивается на измерение интенсивности индивидуальной полосы пропускания излучения. Однако другие пригодные приемники 52 можно сконструировать и из фотодиодов быстрого сканирования, приборов с инжекцией ... |
Еще из этого раздела: 2007901 Устройство для хранения овощей и фруктов 2303347 Способ ведения виноградных кустов 2465767 Оросительный мат для распределения воды на большой площади 2230467 Добавка к пищевым продуктам, биоцидный препарат, 2-(1-окси- 4-гидроксифенилен)-бензохинон (варианты) и способ его получения 2025945 Способ выращивания насаждений сосны 2464784 Защитный слой для растений и деревьев, его изготовление и его применение 2260943 Способ подращивания личинок осетровых рыб 2076583 Способ выращивания растений в теплице и устройство для его осуществления 2056755 Способ регулирования роста овощных культур 2450505 Порционное устройство для вытирания семян трав |
Изобретения в сельском хозяйстве
Обработка почвы в сельском и лесном хозяйствах
Посадка, посев, удобрение
Уборка урожая, жатва
Обработка и хранение продуктов полеводства и садоводства
Садоводство, разведение овощей, цветов, риса, фруктов, винограда, лесное хозяйство
Новые виды растений или способы их выращивания
Производство молочных продуктов
Животноводство, разведение и содержание птицы, рыбы, насекомых, рыбоводство, рыболовство
Поимка, отлов или отпугивание животных
Консервирование туш животных, или растений или их частей
Биоцидная, репеллентная, аттрактантная или регулирующая рост растений активность химических соединений или препаратов
Хлебопекарные печи, машины и прочее оборудование для хлебопечения
Машины или оборудование для приготовления или обработки теста
Обработка муки или теста для выпечки, способы выпечки, мучные изделия
|
|
||
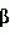 -меркаптоэтанолом и определяют в вышеперечисленных ядерных фракциях протеолитическую и ингибиторную активности. Изобретение позволяет реализовать указанное назначение. 1 табл., 4 ил.
-меркаптоэтанолом и определяют в вышеперечисленных ядерных фракциях протеолитическую и ингибиторную активности. Изобретение позволяет реализовать указанное назначение. 1 табл., 4 ил.