Картофель с повышенным выходом крахмала в пересчете на отдельное растение и способ его полученияПатент на изобретение №: 2289914 Автор: КИСАКА Хирояки (JP), ЯНАГИСАВА Суити (JP), МИВА Тетсуя (JP), АКИЯМА Аи (JP) Патентообладатель: АДЗИНОМОТО КО., ИНК. (JP) Дата публикации: 10 Апреля, 2006 Начало действия патента: 28 Октября, 2004 Адрес для переписки: 129010, Москва, ул. Б.Спасская, 25, стр.3, ООО "Юридическая фирма Городисский и Партнеры", пат.пов. Е.Е.Назиной Изображения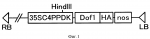  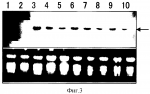 Изобретение относится к биотехнологии, в частности к способу получения картофеля с повышенным содержанием крахмала, а также к таким растениям картофеля. Проводят трансформацию растений картофеля конструкцией, содержащей ген Dof1 кукурузы, кодирующий ДНК-связывающий белок, для экспрессии в них данного гена. Изобретение позволяет получать растения картофеля с повышенным содержанием крахмала. 4 н.п. ф-лы, 3 ил., 2 табл. Область техники Настоящее изобретение относится к картофелю с повышенным выходом крахмала в пересчете на отдельное растение при увеличении урожая картофельных клубней, а также повышении содержания крахмала на единицу массы клубней, к способу получения растений картофеля и к способу получения крахмала из указанных растений картофеля. Предшествующий уровень техники Углеводы откладываются в растениях в основном в форме крахмала. По относительному количеству картофеля, расходуемого в Японии на разные цели, его использование в качестве сырьевого материала для получения крахмала является наивысшим и составляет около 40%, а его использование в пище и в обработанных продуктах питания составляет соответственно 25% и 10%. Потребление крахмала увеличивается, тогда как производство картофеля год от года снижается. В этих условиях для увеличения выхода крахмала необходимо увеличить количество крахмала в картофельных клубнях. С этой целью пытаются улучшить способность усвоения двуокиси углерода, а также способность ассимиляции веществ в запасающем органе с тем, чтобы синтезировать и накапливать избыточное количество сахаридов. Вместе с тем, если даже содержание крахмала в картофельных клубнях удается повысить, количество выращиваемых клубней во многих случаях снижается. Количество крахмала в расчете на отдельное растение можно увеличить лишь в нескольких случаях. Например, Beaujean и соавт. встроили ген С4-фосфоенолпируваткарбоксилазы из сорго (C4-PEPC) и ген НАДФ-зависимой малатдегидрогеназы в растения картофеля с целью повысить эффективность усвоения двуокиси углерода картофелем. Хотя увеличение количества белка, а также увеличение активности встроенных генов было явным, содержание крахмала в клубнях осталось неизменным (ссылка на документ 1, не являющийся патентом). Veramendi и соавт. встроили в растения картофеля ген гексокиназы 1 в антисмысловой ориентации с целью повысить в нем содержание крахмала и достигли снижения гексокиназной активности картофеля до 22% от уровня в природных линиях. В итоге увидели, что содержание крахмала в клубнях не изменилось, тогда как содержание крахмала в тканях листьев увеличилось в три раза (ссылка на непатентный документ 2). Кроме того, при встраивании гена переносчика сахарозы в антисмысловой ориентации, урожай клубней оказался сильно сниженным, хотя содержание крахмала в тканях листьев увеличилось в 5 раз и, таким образом, производство крахмала не увеличилось (ссылка на непатентный документ 3). Таким образом, повысить одновременно содержание крахмала и урожай клубней у картофеля трудно, поэтому сообщений о заметном увеличении крахмала в пересчете на отдельное растение еще не было. [Непатентный документ 1] Beaujean и соавт., Plant Science 160, 1199-1210, 2001. [Непатентный документ 2] Veramendi и соавт., Plant Physiology 121, 123-134, 1999. [Непатентный документ 3] Riesmeier J.W. и соавт., The EMBO J. 13, 1-7, 1994. Описание изобретения Проблема, решаемая в настоящем изобретении Цель настоящего изобретения заключается в разработке способа получения растений картофеля, характеризующихся повышенным содержанием крахмала в пересчете на отдельное растение при увеличении содержания крахмала в картофельных клубнях и увеличении урожая клубней, для увеличения производства крахмала из картофеля. Способы решения данной проблемы Авторы настоящего изобретения считают, что поглощение азота и его метаболизм можно активировать в результате сверхэкспрессии в клетках картофеля транскрипционного регуляторного фактора гена кукурузы Dof1. Транскрипционный регуляторный фактор гена кукурузы Dof1 является положительным регуляторным фактором для гена фосфоенолпируваткарбоксилазы и для гена цитоплазматической дикиназы ортофосфорной кислоты. Считается, что при одновременном увеличении этих ферментативных активностей течение метаболизма углерода сдвигается в сторону системы синтеза аминокислот (Японская Патентная Заявка № 2002-40947). Поэтому ожидается, что если в растении сверхэкспрессируется ген Dof1, то количество синтезируемой в нем аминокислоты увеличивается, и в результате степень азотной ассимиляции повышается. Азот представляет собой наиболее существенный для растений фактор, ограничивающий рост, и предполагается, что когда способность поглощения азота и способность ассимиляции азота у растения повышаются благодаря встраиванию гена Dof1, рост данного растения ускоряется, и увеличенное количество запасных веществ у данного растения может накапливаться. У растений картофеля в период формирования органов накопления запасных питательных веществ, клубней, такие накапливаемые питательные вещества, как полагают, переносятся в клубни путем их перемещения, что способствует формированию клубня и, следовательно, увеличивают урожай клубней. Далее, сахариды, транспортируемые в избыточном количестве, превращаются в крахмал (запасная форма) и накапливаются в большом количестве в этих клубнях. Авторы настоящего изобретения получили трансформированные растения картофеля, в которые встроен ген Dof1, и на фактах подтвердили одновременное увеличение урожая клубней и повышение содержания крахмала на удельный вес клубней. В итоге подтверждено, что количество крахмала, получаемого в отдельном картофельном клубне, возросло. Авторы настоящего изобретения полностью достигли цели, поставленной в настоящем изобретении. А именно, настоящее изобретение относится к растениям картофеля, характеризующимся более высоким урожаем картофельных клубней в расчете на картофельное растение по сравнению с нетрансформированными картофельными растениями, выращиваемыми в тех же условиях, причем ген, кодирующий ДНК-связывающий белок, принадлежащий семейству Dof, встроен в данные растения картофеля. Настоящее изобретение относится к растениям картофеля, характеризующимся более высоким содержанием крахмала в картофельных клубнях по сравнению с нетрансформированными растениями картофеля, выращиваемыми в тех же условиях, причем ген, кодирующий ДНК-связывающий белок, принадлежащий семейству Dof, встроен в данные растения картофеля. Настоящее изобретение относится к растениям картофеля, характеризующимся более высоким содержанием крахмала в пересчете на отдельное растение по сравнению с нетрансформированными растениями картофеля, выращиваемыми в тех же условиях, причем ген, кодирующий ДНК-связывающий белок, принадлежащий семейству Dof, встроен в данные растения картофеля. Настоящее изобретение относится к потомству вышеуказанных растений картофеля, которые несут ген, кодирующий ДНК-связывающий белок, принадлежащий семейству Dof. Настоящее изобретение относится к способу получения растений картофеля, характеризующихся более высоким урожаем клубней в пересчете на отдельное растение, по сравнению с нетрансформированными растениями картофеля, выращиваемыми в тех же условиях, включающему встраивание в растения картофеля гена, кодирующего ДНК-связывающий белок, принадлежащий семейству Dof, для экспрессии данного гена в растениях картофеля. Настоящее изобретение относится к способу получения растений картофеля, характеризующихся более высоким содержанием крахмала в картофельных клубнях, по сравнению с нетрансформированными картофельными растениями, выращиваемыми в тех же самых условиях, включающему встраивание гена, кодирующего ДНК-связывающий белок, принадлежащий семейству Dof, в растения картофеля с целью экспрессии данного гена в растениях картофеля. Настоящее изобретение относится к способу получения растений картофеля, характеризующихся более высоким содержанием крахмала в пересчете на отдельное растение, по сравнению с нетрансформированными растениями картофеля, выращиваемыми в тех же условиях, включающему встраивание гена, кодирующего ДНК-связывающий белок, принадлежащий к семейству Dof, в растения картофеля, которые экспрессируют данный ген. Настоящее изобретение относится к способу получения крахмала, включающему культивирование таких растений картофеля или его потомков и получение крахмала из собранных клубней. Предпочтительное осуществление настоящего изобретения Используемый в настоящем изобретении ДНК-связывающий белок, относящийся к семейству Dof, может представлять собой связывающий ДНК белок, принадлежащий семейству Dof, из другого вида растения, который функционирует в качестве основного контролирующего белка в ходе пополнения углерода либо, иными словами, в качестве группы белков, контролирующих метаболический путь до 2-оксоглутарата, и, в частности, в качестве белка, контролирующего метаболический путь на отрезке от триозы до 2-оксоглутарата в системе гликолиза и последующего цикла трикарбоновых кислот (TCA). Dof1 кукурузы является одним из его предпочтительных примеров. Далее, ДНК-связывающий белок, обладающий делецией, замещением или дополнением одной или нескольких аминокислот в данном ДНК-связывающем белке, принадлежащем Dof-семейству, также используется в настоящем изобретении до тех пор, пока он обладает вышеуказанной функцией. Кроме того, в настоящем изобретении может также использоваться любой ген, способный гибридизоваться в жестких условиях с молекулой нуклеиновой кислоты, кодирующей ДНК-связывающий белок, принадлежащий Dof-семейству. Используемый в данном описании термин "жесткие условия" указывают на условия, в которых образуются так называемые специфичные гибриды. Точно перечислить жесткие условия трудно, потому что они варьируют в зависимости от содержания GC в каждой последовательности, наличия или отсутствия повторяющихся последовательностей и тому подобное. Жесткими условиями являются, например, условия, при которых гибридизуются молекулы нуклеиновой кислоты, обладающие высокой гомологией, к примеру, молекулы нуклеиновых кислот, обладающие, по меньшей мере, 65%-й гомологией, но в которых не гибридизуются нуклеиновые кислоты, обладающие более низкой гомологией. Кроме того, жесткие условия представляют собой такие условия, в которых молекулы нуклеиновых кислот гибридизуются при концентрации соли, соответствующей обычным условиям отмывки при Саузерн-гибридизации (60°С, 1xSSC, 0,1% SDS, предпочтительно 0,1xSSС, 0,1% SDS). Принимая во внимание ту роль, которую играет основной контролирующий ген/белок, а также принимая во внимание тот факт, что Dof1 кукурузы в действительности функционирует также в Arabidopsis thaliana, что раскрыто в данном исследовании, очевидно, что даже когда ДНК-связывающий белок, принадлежащий Dof-семейству и его ген происходят из других видов растений, в предполагаемом растении он обнаруживает функцию, эквивалентную функции Dof1 кукурузы. Используемый здесь термин "обладающий функцией, эквивалентной функции Dof1 кукурузы" свидетельствует о том, что он обладает функцией ускорения транскрипции гена фосфоенолпируваткарбоксилазы и/или гена цитоплазматической дикиназы ортофосфорной кислоты. Вышеуказанный ген из семейства Dof или его кДНК можно сравнительно легко получить следующим образом в соответствии с опубликованной информации о последовательности. Например, что касается гена Dof, основная последовательность его кДНК раскрыта в Genbank под каталожным № Х66076 (SEQ ID NOS 1 и 2). кДНК Dof1 можно сравнительно легко получить путем синтеза ПЦР-праймеров для амплификации ДНК-фракции, содержащей белок - кодирующую часть в соответствии с последовательностью данного белка, и последующего осуществления RT-ПЦР с помощью РНК, экстрагированной из листьев кукурузы в качестве матрицы. В соответствии с настоящим изобретением растения картофеля, характеризующиеся повышенной урожайностью клубней, и/или растения картофеля, характеризующиеся повышенным содержанием крахмала на единицу массу клубней, и/или растения картофеля, характеризующиеся повышенным количеством крахмала в пересчете на отдельное растение, получают после встраивания в растения картофеля конструкции нуклеиновой кислоты, содержащей ген Dof1 кукурузы, и экспрессирования гена Dof1 для получения трансформированных растений картофеля. В соответствии с настоящим изобретением предпочтительно, чтобы средняя масса собранных клубней с растения картофеля превышала, по меньшей мере, в 1,2 раза массу клубней обычного нетрансформированного растения картофеля, выращиваемого в тех же условиях. Что касается содержания крахмала, то предпочтительно, чтобы содержание крахмала на единицу растения у данного картофеля превышало, по меньшей мере, в 1,5 раза содержание крахмала у обычного нетрансформированного растения, выращиваемого в тех же условиях. Используемые в настоящем изобретении конструкции нуклеиновых кислот можно получить способом, хорошо известным в данной области техники. Что касается молекулярных биологических способов выделения конструкции нуклеиновой кислоты и определения ее последовательности, отсылаем к таким печатным материалам как Sambrook и соавт., Molecular cloning - Laboratory manual, the second edition, Cold Spring Harbor Laboratory Press. Если для получения конструкций нуклеиновых кислот, используемых в настоящем изобретении, требуется осуществить генную амплификацию, например, с помощью способа ПЦР, то ее методики раскрыты, например, у F.M.Ausubel и соавт. (eds.), Current Protocols in Molecular Biology, John Wiley & Sons, Inc. (1994). Используемые в настоящем изобретении конструкции нуклеиновых кислот, как правило, могут содержать соответствующие промоторы, функционирующие в растительных клетках, такие как гены нопалинсинтазы, промотора 35S вируса мозаики цветной капусты (CaMV35S), соответствующий терминатор, такой как терминатор гена нопалинсинтазы, другие последовательности, необходимые для успешной экспрессии, и маркерные гены для отбора трансформанта, такие как гены устойчивости к химическим веществам, например, гены устойчивости к канамицину, устойчивости к G418 и к гигромицину. Используемый для конструкций нуклеиновых кислот промотор может представлять собой конститутивный промотор, либо органоспецифичный или специфичный для стадии роста промотор. Промотор можно выбрать в зависимости от используемого хозяина, необходимого уровня экспрессии, органа, в котором, в частности, предполагается экспрессия, или от стадии роста. В предпочтительном варианте осуществления настоящего изобретения используют сильный промотор, который неспецифически экспрессируется в органе или на стадии роста. Примером таких промоторов является CaMV35S-промотор. Что же касается органоспецифичных промоторов, то используют промотор гена фазеолина, промотор гена пататина и им подобные. В более предпочтительном варианте осуществления настоящего изобретения используют конструкцию, в которую вставлен ген Dof1 под сильным конститутивным промотором, таким как CaMV35S. Способ встраивания генов в настоящем изобретении особенно не ограничен. Способ встраивания генов в растительные клетки или в растения, известный в данной области техники, можно выбрать в зависимости от определенного хозяина. Например, в варианте осуществления настоящего изобретения используют способ встраивания гена с помощью Agrobacterium. В такой системе трансформации желательно использовать бинарный вектор. При использовании Agrobacterium применяемая для трансформации конструкция нуклеиновой кислоты дополнительно содержит Т-участок, примыкающий к ДНК-последовательности, которую встраивают в растительные клетки. В предпочтительном варианте осуществления настоящего изобретения вводимую последовательность встраивают между граничащих слева и справа последовательностей Т-ДНК. Подходящее конструирование трансформирующего вектора, основанного на такой Т-ДНК, хорошо известны специалистам в данной области техники. Условия инфицирования растения с помощью Agrobacterium, обладающей такой конструкцией нуклеиновой кислоты, хорошо известны специалистам в данной области техники. Что касается методов и условий, отсылаем к "Model Shokubutsu no Jikken Protocol (Experimental protocol of model plants), Edition of Rice plant and Arabidopsis thaliana" (a separate volume of Saibo Kokaku) edited by Shujun-sha in 1996. В настоящем изобретении можно также использовать и другой способ встраивания гена. Примеры способов встраивания гена, которые можно использовать здесь, включают, например, способ встраивания ДНК в протопласты с помощью полиэтиленгликоля или кальция, способ трансформации протопластов путем электропорации и способ встраивания с помощью бомбардирующих частиц. Обработанные таким образом растительные клетки или им подобные отбирают для трансформации. Данный отбор можно осуществить в соответствии с экспрессией маркерного гена, присутствующего в данных конструкциях нуклеиновых кислот, используемых для трансформации. Например, если маркерные гены устойчивы к лекарственным средствам, их можно отобрать путем культивирования или путем выращивания растительных клеток в среде, содержащей соответствующую концентрацию антибиотика, гербицида или подобного вещества. Если данные маркерные гены представляют собой гены Полученный таким образом растительный организм можно выращивать с помощью обычного способа в тех же условиях, в которых выращивают нетрансформируемое растение. Для идентификации трансформированного растения, содержащего конструкцию нуклеиновой кислоты настоящего изобретения, можно использовать различные молекулярно-биологические методики в дополнение к вышеописанной селекции, основанной на маркерном гене. Например, для детектирования фракции встроенной рекомбинантной ДНК и ее структуры можно использовать Саузерн-гибридизацию или ПЦР. Нозерн-гибридизацию или RT-ПЦР можно использовать для детектирования и определения продуктов транскрипции РНК из встроенных конструкций нуклеиновых кислот. Экспрессию гена Dof1 в полученном трансформанте оценивают путем определения количества белка Dof1 или количества мРНК. Например, количество Dof1-белка можно определить способом Вестерн-блоттирования или подобным образом, а количество мРНК можно определить способом Нозерн-блоттирования или количественным RT-ПЦР-способом. После подтверждения экспрессии гена Dof1 в трансформированных таким образом растениях картофеля определяют урожай клубней и содержание крахмала в данных клубнях. Трансформированные растения картофеля выращивают в обычных условиях для почвенной культуры. Растительные организмы, используемые для данной почвенной культуры, представляют собой саженцы, полученные в результате адаптации трансформированных, культивируемых продуктов или микроклубней, полученных с помощью тканевой культуры. Клубни, полученные с помощью обычной культуры, оценивают на основе числа клубней, собранных у отдельного растения, и его общей массы. Содержание крахмала в клубнях можно определить путем измельчения целых клубней, получения из них экстракта и определения содержания крахмала, например, ферментативным способом. Получение растительного экстракта и определение содержания крахмала можно осуществить, например, способом Agarie и соавт. (S.Agarie, Plant Science, 2002, 162, 257-265). Вес и содержание крахмала в собранных клубнях определяют, если значимое различие составляет 5% или меньше в таком статистическом тесте, как t-критерий. Последовательность из конструкций нуклеиновой кислоты в интродуцированном гене может быть либо гемизиготной, либо гомозиготной в трансформированном растении. Посевной картофель на практике размножают вегетативно, а введенный интродуцированный ген устойчиво передается в потомстве безотносительно от способа встраивания в данную последовательность. Посадочный картофель полученный в результате трансформированных растений картофеля можно размножить и сохранять обычным способом, хорошо известным в данной области техники. Крахмал можно получить из клубней трансформированных растений картофеля обычным способом, хорошо известным специалистам в данной области техники. Примеры Настоящее изобретение иллюстрируется особо нижеследующими примерами получения растений с помощью воздействия повышенной экспрессии гена Dof1, анализа свободных аминокислот, изучения урожайности и определения содержания крахмала. Пример 1. Интеграция кукурузного гена Dof1 в растительный трансформирующий вектор Ген Dof1 интегрирован из плазмиды 35SC4PPDK-Dof1-HA (The Plant Cell 1998, 10 (Jan), 75-89) в растительный трансформирующий вектор pBI121 (Clontech Co.), как описано ниже. Плазмида 35SC4PPDK-Dof1-HA обладает участками ДНК, в которой область, простирающаяся от TATA-бокса до сайта инициации трансляции кукурузного дикиназного гена С4-пируватортофосфорной кислоты, и кДНК Dof1 объединяют ниже от энхансера 35S вируса мозаики цветной капусты, а 2 молекулы кДНК фитогемагглютинина последовательно располагают правее в качестве эпитопной метки и, наконец, присоединяют сайт терминации нопалинсинтазы. Указанные участки ДНК можно выделить двойным расщеплением ферментами рестрикции XhoI и EcoRI. В связи с этим предпринята попытка интегрировать участок XhoI-EcoRI в Т-ДНК-область pBI121. Вначале расщепляли pBI121 по HindIII и его конец затупляли с помощью клонированной ДНК-полимеразой Pfu от Stratagene Co. После связывания с фосфорилированным XhoI-линкером (Takara Shuzo Co., Ltd.) с помощью Т4-ДНК-лигазы (Takara Shuzo Co., Ltd.) с последующим расщеплением по XhoI, линкерные фракции удаляли с помощью гель-фильтрации на колонке MicroSpin S-300 от Amersham-Pharmacia Co. После автоциклизации с помощью Т4-ДНК-лигазы получали плазмиду, обладающую сайтом XhoI вместо сайта HindIII. Данная плазмида содержала в качестве уникальных сайтов XhoI и EcoRI. Полученную плазмиду расщепляли по XhoI и EcoRI, а затем фракцию XhoI-EcoRI, отдельно полученную из 35SC4PPDK-Dof1-HA, интегрировали в это место с образованием pBI121Dof1 (фиг. 1). Пример 2. Получение трансформантов картофеля Растения картофеля (сорт: May Queen) трансформировали по способу Gordon и соавт. (Plant Cell Reports, 1993, 12:324-327). Стерильные растения, полученные путем выращивания конуса нарастания, укореняли в MS-жидкую среду. В нее добавляли 16%-й раствор сахарозы. Стимулировали микроклубни путем их культивирования в темном месте. Микроклубни, стимулированные в стерильных условиях, обдирали, резали на тонкие диски (микроклубневые диски) и использовали для инфицирования Agrobacterium. Полученные микроклубневые диски рекультивировали в среде для повторной дифференцировки (неорганические соли MS, витамин В5, 3% сахарозы, 2 мг/л зеатина, 0,1 мг/л индолацетата, 2,5 мг/л gellan gum, рН 5,8) в течение 24 часов. Плазмиду pBI121 встраивали в Agrobacterium C58C1Rif путем тройного скрещивания с E. coli, несущей pBI121Dof1, и с хелперной E. coli HB101/pRK203. Agrobacterium, содержащая данный сконструированный ген, инокулировали в YEP-среду (10 г/л бактотриптона, 10 г/л дрожжевого экстракта и 1 г/л глюкозы), содержащую 50 мг/л канамицина, и культивировали со встряхиванием при 28°С в течение 24 часов. Полученный культуральный продукт погружали в разбавленную смесь суспензии Agrobacterium и культуральной среды (неорганическая соль MS, витамин В5, 3% сахарозы, 2 мг/л зеатина, 0,1 мг/л индолацетата, 1,0 мг/л 3,5-диметокси-4'-гидроксиацетофенона, Aldrich Co., рН 5,8) и оставляли стоять в течение 10 минут. Затем излишек культуральной жидкости Agrobacterium вытирали стерильной фильтровальной бумагой, и данный продукт переносили в культуральную среду, используемую для прекультивирования. После сокультивирования при 26°С в течение 48 часов (длина дня: 16 часов) их переносили в селективную среду (неорганические соли MS, витамин В5, 3% сахарозы, 2 мг/л зеатина, 0,1 мг/л индолацетата, 50 мг/л канамицина, 300 мг/л натриевой соли цефотаксима и 2,5 мг/л gellan gum, рН 5,8) и продолжали культивирование в тех же условиях. Этот продукт переносили в новую селективную среду каждые две недели и одновременно повторно дифференцированные проростки переносили в среду для укоренения (неорганические соли MS, витамин В5, 3% сахарозы, 50 мг/л канамицина, 100 мг/л натриевой соли цефотаксима и 2,5 мг/л gellan gum, рН 5,8). Нарастающий конус укорененного растения переносили в новую среду для укоренения с повторением теста на устойчивость 3 или 4 раза. Отбирали 8 корешков картофеля, трансформированного Dof1, обладающего устойчивостью к канамицину. Пример 3. Подтверждение встраивания гена ДНК экстрагировали из 8 отобранных растений картофеля, устойчивых к канамицину, и из нетрансформированных растений картофеля. Способ экстракции ДНК представлял собой способ Honda и соавт. (Honda and Hirai, 1990, Jpn Breed 40:339-348). ПЦР-анализ осуществляли с использованием экстрагированной ДНК и праймеров к промоторной части, встроенной в ген Dof1, и части терминатора: 5'-TTCCATTGCC CAGCTATCTG TCACTT-3' (SEQ ID NO.3) и 5'-TCATCGCAAG ACCGGCAACA GGATTC-3' (SEQ ID NO.4) а также праймеров для амплификации гена NPTII в векторе: 5'-CCC CTC GGT ATC CAA TTA GAG-3' (SEQ ID NO.5) и 5'-CGG GGG GTG GGC GAA GAA CTC CAG-3' (SEQ ID NO.6). Условия данной реакции были следующими: 94°С - 3 минуты; 94°С - 45 секунд, 55°С - 30 секунд, 72°С - 90 секунд, 35 циклов; 72°С - 10 минут. Для осуществления данной реакции использовали прибор PCR system 2400 of Perkin-Elmer Co. Полученный ПЦР-продукт подвергали электрофорезу в 1%-м агарозном геле (буфер TAE) и окрашивали этидийбромидом. В итоге, в отобранных трансформированных растениях картофеля оценивали размер полосы, содержащей Dof1-ген (около 1,5 т.п.н., фиг. 2а) и полосы, специфичной для NPTII-гена (около 1,1 т.п.н., фиг. 2b). Данные полосы не распознавались в нетрансформированных растениях картофеля. Из этих данных понятно, что ген Dof1, содержащий TDHA-домен, встроен в данные трансформированные растения картофеля. Пример 4. Подтверждение экспрессии встроенного гена (Нозерн-анализ) 8 корешков трансформированных растений картофеля, для которых подтверждено встраивание гена Dof1, подвергали Нозерн-анализу для подтверждения экспрессии указанного встроенного гена. Тотальную РНК экстрагировали из ткани листьев трансформированных растений картофеля, через один месяц после их посадки в почву, с помощью RNeasy Plant mini Kit of Qiagen Co. 10 мкг экстрагированной РНК подвергали электрофорезу в 1,2%-м агарозном геле, содержащем 18% формальдегида, и затем окрашивали этидийбромидом. После блоттирования на нейлоновую мембрану (HybondN+) полученный продукт фиксировали под УФ и гибридизовали. Полноразмерный ген Dof1 использовали в качестве зонда. Нозерн-блоттирование и получение зонда осуществляли с помощью наборов DIG-High Prime DNA Labeling и Detection Starter Kit II, а также PCR DIG Probe Synthesis Kit of Roche Diagnostics Co. В итоге, в листовой ткани трансформированных растений картофеля подтверждали наличие полосы, специфичной для гена Dof1 (фиг. 3). Установлено, что встроенный ген Dof1 транскрибируется и экспрессируется в ткани листьев трансформированных растений картофеля. В нетрансформированных растениях картофеля не обнаруживали полосу, специфичную для гена Dof1. Пример 5. Анализ урожайности трансформированных растений картофеля Анализировали урожайность трансформированных растений картофеля. Для проверки использовали стерильные растения из числа трансформированных растений картофеля и нетрансформированные растения картофеля. После акклиматизации 0,5 кг плодородной земли (общее содержание азота 0,2 г) загружали в цветочный горшок (3,5 л), после чего в него доверху вносили вермикулит (непитательная среда). Через два месяца после начала выращивания 0,3 кг (общее содержание азота 0,12 г) плодородной почвы использовали в качестве почвенного покрова. В период выращивания число стебельков сокращалось до одного. На третий месяц после пересадки растения картофеля собирали и взвешивали надземную часть, взвешивали клубни и подсчитывали количество клубней. С помощью данного анализа установлено, что по сравнению с нетрансформированными растениями картофеля растения картофеля, трансформированные Dof1, обладали в 1,24-1,50 раза большей массой надземной части, в 1,05-1,40 раз большей массой клубней и в 1,42-1,90 раз большим количеством клубней (Таблица 1). Таблица 1.Результаты анализа урожайности растений картофеля, трансформированных Dof1 n=3 Вес надземной части (г)Общий вес клубней (г) Общее количество клубнейНетрансфомируемое растение31,02±3,66 96,32±1,606,7±0,8 Dof1-141,85±4,47 122,80±11,4812,7±1,0 Dof1-238,56±3,49 100,98±8,399,7±0,4 Dof1-345,04±0,33 132,29±0,9610,5±0,2 Dof1-446,38±0,33 132,35±1,9512,3±1,0 Dof1-539,39±2,77 109,53±9,5110,7±1,3 Dof1-639,61±1,42 120,78±13,289,5±1,2 Dof1-841,83±1,76 129,02±12,3012,3±0,2 Dof1-942,43±0,62 135,23±4,2712,3±0,5Пример 6. Определение содержания крахмала в клубнях Клубни трансформированных растений картофеля и нетрансформированных растений картофеля замораживали (-80°С) сразу после их сбора и держали в замороженном состоянии, а также определяли в них содержание крахмала. В каждой группе использовали два клубня. Данные целые клубни опрыскивали жидким азотом. Экстрагирование осуществляли способом S.Agarie и соавт. (Plant Science 162, 2002, 257-265). Содержание крахмала определяли с помощью набора F-kit starch of Roche Diagnostics Co. В итоге, содержание крахмала в клубнях растений картофеля, трансформированных Dof1, оказалось увеличенным с 1,06 до 1,53 раза по сравнению с клубнями нетрансформированных растений картофеля. Содержание крахмала в пересчете на каждое картофельное растение оказалось увеличенным в 2 раза или даже больше по сравнению с нетрансформированным картофельным растением (Таблица 2). Таблица 2.Содержание крахмала в клубнях растений картофеля, трансформированных Dof1 Содержание крахмала (мкмоль/г.F.W.)Вес клубня (г) Общее содержание крахмала (мкмоль)соотношение Нетрансформант 35,71±4,2696,32 3439,521,0 Dof1-148.80±3,60 122,805993,031,7 Dof1-337,80±3,14 132,295001,13 1,5Dof1-4 42,12±2,61132,25 5570,851,6 Dof1-954,91±0,05 135,237425,482,2Аминокислотный анализ тканей листьев и стеблей растения, выращенного в стерильных условиях, осуществляли, чтобы установить, что общее содержание аминокислот у растений картофеля, трансформированных Dof1, выше, чем в нетрансформированных растениях картофеля в 1,88 раза или меньше. <Таблица последовательностей без текста> SEQ ID NO.1; Последовательность оснований кДНК кукурузного гена Dof1 SEQ ID NO.2; Последовательность аминокислот, кодируемая кДНК гена Dof1 кукурузы SEQ ID NOS.3 и 4; Праймеры в промоторной и терминаторной части, в которые встроен ген Dof1 SEQ ID NOS.5 и 6; Праймеры для амплификации гена NPTII в данном векторе. Краткое описание чертежей: На фиг. 1 схематически изображена встроенная часть Dof1 в плазмиде pBI121Dof1. На фиг. 2 представлены результаты ПЦР-анализа растений картофеля, трансформированных Dof1, где: (А) представлены результаты ПЦР-реакции с праймером, содержащим Dof1-ген, (В) представлены результаты ПЦР-реакции с праймером, специфичным для гена NPTII. Дорожка 1 представляет нетрансформант; дорожка 2 представляет Dof1-1; дорожка 3 представляет Dof1-2; дорожка 4 представляет Dof1-3; дорожка 5 представляет Dof1-4; дорожка 6 представляет Dof1-5; дорожка 7 представляет Dof1-6; дорожка 8 представляет Dof1-8; и дорожка 9 представляет Dof1-9. На фиг. 3 представлены результаты Нозерн-анализа листовой ткани картофельных растений, трансформированных Dof1. Дорожки 1-2 представляют нетрансформант; дорожка 3 представляет Dof1-1; дорожка 4 представляет Dof1-2; дорожка 5 представляет Dof1-3; дорожка 6 представляет Dof1-4; дорожка 7 представляет Dof1-5; дорожка 8 представляет Dof1-6; дорожка 9 представляет Dof1-8; и дорожка 10 представляет Dof1-9. СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ Формула изобретения1. Растение картофеля, характеризующееся более высоким содержанием крахмала в картофельных клубнях по сравнению с нетрансформированными растениями картофеля, выращиваемыми в тех же условиях, где в указанное растение картофеля встраивают ген Dof1 кукурузы, кодирующий белок, связывающий ДНК. 2. Потомство растения картофеля по п.1, которое несет ген Dof1 кукурузы, кодирующий белок, связывающий ДНК. 3. Способ получения растения картофеля, характеризующегося более высоким содержанием крахмала в картофельных клубнях по сравнению с нетрансформированными растениями картофеля, выращиваемыми в тех же условиях, предусматривающий встраивание гена Dof1 кукурузы, кодирующего белок, связывающий ДНК, в растение картофеля для экспрессии в нем данного гена. 4. Способ получения крахмала, предусматривающий выращивание растения картофеля по п.1 или потомства по п.3 и получение крахмала из выращенных клубней. Популярные патенты: 2175189 Способ регенерации растений сорго в культуре in vitro ... среде, содержащей аминокислоты L-аспарагин (1.0 г/л) и L-пролин (2.0 г/л), значительным образом стимулировало рост эмбриогенного каллуса сорго. Зависимость массы ЭК от концентрации ионов NO3 - и NH4 + при одновременном увеличении их содержания в питательной среде, как было описано выше, графически представлена на чертеже (А и Б соответственно). Учитывая, что в прототипе концентрация ионов NO3 -составляет 39.9 mM, а также величину ошибки средней (99 мг), очевидно, что статистически значимый эффект наблюдается в диапазоне концентраций 61-88 mM NO3 -. Аналогичным образом, учитывая, что в прототипе концентрация ионов NH4 + составляет 20.6 mM, статистически значимый эффект ... 2084132 Устройство для выращивания растений ... непроницаемый для корней материал 18. Концы пленочного эластичного материала 17 и пленочного пористого непроницаемого для корней материала 18 спущены в питательные каналы 7. Питательный раствор из нижнего бака 20 насосом 21 с приводом 22 подают по трубопроводу в средний питательный канал 7. Питательный раствор, уровень которого поднимается выше краев среднего канала 7 растильни 1 перетекает по плоским капиллярам между пленочным пористым материалом 16 и пленочным пористым непроницаемым для корней материалом 18 через патрубки 11, 12, 13 каналов 7 по трубопроводам 38, 39 в нижний бак питательного раствора 20. Пленочный пористый эластичный материал 17, уложенный вместе с вкладышем ... 2395497 Способ стимулирования роста подсолнечника регулятором роста ... - М.: Агропромиздат, 1987. - С.33-80].Задачей настоящего изобретения является расширение арсенала биологически активных веществ, полученных синтетическим путем, для применения их в сельском хозяйстве в качестве регуляторов роста растений.Это достигается применением О-(замещенный фенил)карбонил-4,6-диметил-2-хлорпиридил-3-амидоксимов 1-2 путем предпосевной обработки семян подсолнечника. При этом O-(замещенный фенил)карбонил-4,6-диметил-2-хлорпиридил-3-амидоксимы 1-2 получены известным способом - ацилированием 4,6-диметил-2-хлорпиридил-3-амидоксима 4 хлорангидридами ароматических кислот в среде безводного бензола в присутствии триэтиламина в качестве акцептора хлороводорода [1]: В ... 2472951 Машина (варианты) ... машины, повышает безопасность работы в холодной зоне: облегчает обработку земли зимой и ремонт машин летом.Перекладки поршней около мертвых точек, сил давления на боковые поверхности, нарушения форм /цилиндричности/ их и цилиндров из-за неравномерности износа, наличие шатунов, колен вала часто с подшипниками скольжения, изменение плеча сил/крутящего момента, необходимость непрерывной смазки, высокие требования к маслам, металлам, температурам снижает ресурс и работоспособность.Нить с шариками или рычагами может преобразовать импульсы во вращение двух конусов сотового вариатора /фиг.5/ регулятора передаточного числа силовой передачи. Нитевой привод 21 к колесам как ... 2469534 Перезаряжаемая электронная ловушка для животных с перегородкой, механическим переключателем в конфигурации с множеством поражающих пластин ... пространство между своим нижним краем и дном корпуса, чтобы вынудить уничтожаемое животное сжиматься под перегородкой, затрудняя для животного возврат назад, когда оно соприкасается со вторым электродом. Еще одной задачей настоящего изобретения является создание электронной ловушки для животных, в которой переключатель помещен перед перегородкой, которая имеет свободный край, перекрывающий средний электрод для минимизации возможности ускользания грызуна. Еще одной задачей настоящего изобретения является создание электронно-управляемой, механически синхронизированной ловушки для животных, которая обеспечивает обнаружение отсутствия животного после подачи высокого напряжения и затем ... |
Еще из этого раздела: 2494593 Способ повышения селена в чесноке горной зоны 2088063 Широкозахватный сельскохозяйственный агрегат 2231250 Устройство для промышленного выращивания земляники и других растений 2178965 Картофелекопатель ручной мотыжный 2053661 Устройство для сколачивания ульевых рамок 2490849 Способ переработки безподстилочного помета птиц клеточного содержания и навоза свиней в топливные брикеты 2236787 Способ испытаний опрыскивателей и устройство для его осуществления 2409937 Растение с высоким содержанием ребаудиозида а 2108700 Способ оценки горных сенокосов и пастбищ 2451442 Способ обогащения селеном овощей и злаков |
Изобретения в сельском хозяйстве
Обработка почвы в сельском и лесном хозяйствах
Посадка, посев, удобрение
Уборка урожая, жатва
Обработка и хранение продуктов полеводства и садоводства
Садоводство, разведение овощей, цветов, риса, фруктов, винограда, лесное хозяйство
Новые виды растений или способы их выращивания
Производство молочных продуктов
Животноводство, разведение и содержание птицы, рыбы, насекомых, рыбоводство, рыболовство
Поимка, отлов или отпугивание животных
Консервирование туш животных, или растений или их частей
Биоцидная, репеллентная, аттрактантная или регулирующая рост растений активность химических соединений или препаратов
Хлебопекарные печи, машины и прочее оборудование для хлебопечения
Машины или оборудование для приготовления или обработки теста
Обработка муки или теста для выпечки, способы выпечки, мучные изделия
|
|
||

 -глюкоронидазы или гены люциферазы, их активность скринируют для отбора трансформанта. Если идентифицированный таким образом данный трансформант представляет собой не растительный организм, а протопласт, каллус, эксплантат или им подобные, его регенерируют в данный растительный организм. Такую регенерацию можно осуществить способом, выбранным в зависимости от растения-хозяина, используя способы, известные в данной области техники.
-глюкоронидазы или гены люциферазы, их активность скринируют для отбора трансформанта. Если идентифицированный таким образом данный трансформант представляет собой не растительный организм, а протопласт, каллус, эксплантат или им подобные, его регенерируют в данный растительный организм. Такую регенерацию можно осуществить способом, выбранным в зависимости от растения-хозяина, используя способы, известные в данной области техники.




