Способ получения белковых веществПатент на изобретение №: 2250264 Автор: РЕСКИ Ральф (DE), ГОРР Гильберт (DE) Патентообладатель: ГРИНОВЕЙШН БАЙОТЕК ГМБХ (DE) Дата публикации: 10 Марта, 2004 Начало действия патента: 27 Сентября, 2000 Адрес для переписки: 129010, Москва, ул. Б.Спасская, 25, стр.3, ООО "Юридическая фирма Городисский и Партнеры", пат.пов. Е.Е.Назиной Изобретение относится к области биотехнологии. Белковые вещества получают не переработкой растительной ткани, а из среды, в которой они культивируются. Использование в качестве растительной ткани протонемы мха позволяет выделять из нее гетерологичные белки в их активной форме, не разрушая продуцирующие ткани и клетки. 5 з.п. ф-лы. Настоящее изобретение в целом относится к получению белковых веществ из растительного субстрата. В частности, настоящее изобретение относится к новому способу получения желаемых белковых веществ из мхов. Использование биотехнологических методов в производственных целях дает человеку возможность получать вещества, которые не могут быть получены экономическими или другими путями, например химическим синтезом, и которые не доступны в природе в количестве, достаточном для взаимодействия в качестве сырья. Хотя известно более 10000 вторичных метаболитов растений, с помощью культур растительных клеток на промышленном уровне получают только несколько таких соединений. Эти соединения являются преимущественно фармацевтически активными вторичными метаболитами. Примерами, которые могут быть приведены, являются а) берберин (объем производства составляет 4000 л), который обладает бактериостатическим и фунгицидным действием (Y. Fujita и М. Tabata в: Plant tissue and cell culture, plant science; том 3, стр. 169, C.E.Green et al. (Ed.), A.R.Liss Inc., Нью-Йорк (1987)), b) шиконин (объем производства 750 л), который действует как антибиотик и обладает противовоспалительной активностью (М. Tabata и Y. Fujita в: Biotechnology in plant science; стр. 207-218, P. Day et al. (Ed.), Academic Press, Орландо (1985)) и с) паклитаксел (объем производства 75000 л), более широко известный под названием таксол, который имеет противоопухолевую активность (М.Jaziri et al., Taxus sp. cell, tissue and organ cultures as alternative sources for taxoids production: a literature survey, Plant Cell Tiss. Org. Cult., 46, стр. 59-75 (1996)). Более важным биотехнологическим методом, в котором используются культуры растительных клеток, является биотрансформация дигитоксина в дигоксин, сердечный и циркуляторный лекарственный препарат. Такую стереоспецифическую реакцию гидроксилирования успешно проводят в биореакторной культуре Digitalis lanata (E.Reinhard и W.Kreis, Kultivierung von pflanzlichen Zeilen im Bioreaktor [Plant cell culture in the bioreactor], Bio. Engin., 5, стр. 135-136 (1989)) с высоким выходом. Современные подробные обзоры по использованию культур растительных клеток в биотехнологии могут быть найдены у Н.-Р. Muhlbach, Use of plant cell cultures in biotechnology, Biotecnnol. Annu. Rev., 4, стр. 113-176 (1998). Разработка методов генетической трансформации высших растений в начале восьмидесятых позволила значительно увеличить продуктивность растений для специфических вторичных конструкций путем трансформации генов специфических ключевых ферментов рассматриваемых метаболических путей. Использовались не только трансгенные интактные растения, но также культуры растительных клеток. Примеры, которые могут быть приведены, включают сверхэкспрессию бактериальной лизиндекарбоксилазы в трансгенной культуре корневых волосков Nicotiana tabacum, которая увеличивает выход биогенных аминов кадаверина и анабазина до 14 раз (J. Berlin et al., Genetic modification of plant secondary metabolism: Alteration of product levels by overexpression of amino acid decarboxylases, в: Advances in Plant Biology, Studies in Plant Science,, том 4, стр. 57-81, D.D.Y. Ryu и S. Furasaki (Ed.), Elsevier, Амстердам (1994)). Однако возможность переноса ДНК в растения не только делает возможными количественные и качественные изменения растительных конструкций; кроме того, растения и культуры растительных клеток становятся более интересными для получения гетерологичных белков (A.S.Moffat, High-Tech plants promise a bumper crop of new products. Science 256, стр. 770-771 (1992)), за основу были взяты два различных подхода. Первый подход включает в себя получение гетерологичных белков из трансгенных интактных растений. Кроме получения антител из трансгенных табачных растений (J.K.-C.Ma et аl., Generation and assembly of secretory antibodies in plants, Science 268, стр. 716-719 (1995)) были описаны экспрессия и точный процессинг альбумина сыворотки человека как в трансгенных растениях табака, так и в трансгенных растениях картофеля (Р.С.Sijmons et al., Production of correctly processed human serum albumin in transgenic plants, Bio/Techn., 8, стр. 217-221 (1990)). В трансгенных растениях табака экспрессировался также эпидермальный фактор роста человека (hEGF) (A.-H.Salmanian et al., Synthesis and expression of the gene for human epidermal growth factor in transgenic potato plants, Biotechnol. Lett., 18, стр. 1095-1098 (1996)). Впрочем, также использовали и другие растения. Используя Arabidopsis thaliana и Brassica napus, успешно получали Leu-энцефалин (Е. Krebbers и J. Vandekerckhove, Production of peptides in plant seeds, Tibtech., 8, стр. 1-3 (1990)). Кроме того, для экспрессии химерных вирусных частиц, которые действуют как вакцины, использовали трансгенные растения Vigna unguiculata (К.Palsgaard et al., Plant-derived vaccine protects target animals against a viral disease, Nat. Biotech., 15, стр. 248-252 (1997)). Основным неудобством при работе с интактными растениями, в случаях, описанных выше в качестве примера, является необходимость их выращивания, что отнимает много времени и является дорогостоящим, и большой объем культивирования, необходимый для производства в промышленных объемах. Кроме того, для выделения желаемого целевого вещества из интактных растений обычно требуется сложный многостадийный процесс, в особенности, когда стабильность и качество продуктов должны соответствовать высоким требованиям, как в случае с веществами, используемыми для фармацевтических и пищевых целей. Во втором подходе для получения антител используются культуры трансгенных клеток табака. Описаны, например, экспрессия антител и их секреция в среду (N.S. Magnuson et al., Enhanced recovery of a secreted mammalian protein from suspension culture of genetically modified tobacco cells, Prot. Expr. Pur., 7, стр. 220-228 (1996)). Так как гетерологичные белки сложно выделить из клеток, секреция целевого белка в среду является значительным усовершенствованием. Кроме того, продукция рекомбинантных фармацевтически важных белков в культуру клеток также вызывает интерес с точки зрения сохранности, так как трансгенные растительные клетки можно выращивать исключительно в биореакторах и их не нужно выделять. Получение необходимой массовой культуры возможно благодаря разработке биореакторов для культур гетеротрофных растительных клеток в крупных масштабах (например, M.L.Shuler et al., Bioreactor engineering as an enabling technology to tap biodiversity: The case of taxol., Ann. N.Y. Acad. Sci., 745, стр. 455-461 (1994)). Основным неудобством второго подхода, в котором используются растительные суспензионные культуры, является низкая скорость роста, относительно низкое образование вторичных метаболитов, ингибирование образования продукта при высокой плотности клеток и, как следствие, низкая продуктивность на объем, образование агрегатов и компонентов клеточной стенки и возрастание чувствительности клеток к поперечному сдвигу. Также необходимо принять во внимание тот факт, что при использовании гетеротрофных клеточных культур всегда должна использоваться сложная среда с разнообразными компонентами, некоторые из которых являются дорогостоящими. Однако самым серьезным неудобством, о котором следует упомянуть, является наличие сомаклональных вариаций в клеточных культурах растений in vitro, которые вызывают количественные и качественные изменения при получении гетерологичных белков (см., например, M.G.K. Jones и К. Lindsey, Plant biotechnology, в: Molecular biology and biotechnology, J.M.Walker и E.W.Gingold (Eds.), 2nd Ed., Royal Soc. of Сhеm., Burlington House, Лондон (1988)). Гетерогенность образующихся продуктов и их функциональных свойств недопустима, в особенности, в связи с производством фармацевтических препаратов и других желаемых веществ, для официального одобрения которых требуется надежная гарантия качества и стандартизация способа производства. Целью настоящего изобретения, таким образом, является способ для стандартизированного производства гетерологичных белковых веществ из растительного субстрата, который существенным образом устранит не только вышеописанные трудности использования интактных растений, но также трудности использования клеточных культурных систем. В соответствии с изобретением цель достигается разработкой нового способа получения гетерологичных белковых веществ из растительного субстрата, согласно которому полностью дифференцированные мхи выращивают при стандартных условиях и полученные белковые вещества выделяют из культуральной среды по существу без разрушения продуцирующих тканей или клеток. Как здесь используется, термин “белковые вещества” охватывает пептиды, полипептиды и белки, а также их фрагменты, которые являются подходящими, в частности, для диагностических, фармацевтических и пищевых целей. Также он охватывает молекулы, которые содержат пептидные связи и которые транслируются растительным субстратом. В предпочтительном осуществлении по настоящему изобретению желательные гетерологичные белковые вещества высвобождаются в культуральную среду в своей биологически активной форме. Как используется в настоящем описании, термин “биологически активные” означает, что целевые вещества с таким признаком имеют желаемые или требуемые функциональные свойства для соответствующей цели. Например, если желательно получение антител, то полученные белки или их функциональные фрагменты являются биологически активными, если они способны осуществлять ожидаемое специфическое связывание с антигеном. Специалисту в данной области очевидно, что для таких целей не всегда требуется нативный белок, а можно найти эпитопы или низкомолекулярные структуры, которые обеспечивают желаемую биологическую активность или действие. Например, фермент является биологически активным, если он способен преобразовывать свой целевой субстрат. В другом предпочтительном воплощении изобретения в культуральной среде, которая по существу не содержит cахаров, витаминов и фитогормонов или их функциональных фрагментов, выращивают растительный субстрат в виде интактных мхов. Способ по изобретению позволяет выращивать интактные и полностью дифференцированные растения в фотоавтотрофных условиях, которые могут быть стандартизированы, то есть не требующих добавления cахаров, витаминов, фитогормонов и тому подобное, как это требуется для предшествующей гетеротрофной суспензионной клеточной системы. Так как используется недорогая и простая питательная среда, стадии получения и очищения желаемых целевых веществ значительно облегчены. В способе по изобретению используемым растительным субстратом предпочтительно является интактный мох, выбранный из группы мхов, включая печеночники, с видами родов Physcomitrella, Funaria, Sphagnum и Ceratodon, а также Marchantia и Sphaerocarpos, которые являются особенно предпочтительными для использования. Способ по настоящему изобретению наиболее предпочтительно проводить с использованием мха Physcomitrella patens. В следующем предпочтительном воплощении конструкция нуклеиновой кислоты, используемая для трансформации, кодирует не только желаемое белковое вещество, а и транспортный белок для секреции вещества из клетки-хозяина в культуральную среду. В соответствии с изобретением можно использовать любую аутологичную и гетерологичную последовательность нуклеиновой кислоты, известную специалисту в данной области, и ее можно использовать для получения экспрессионной кассеты для трансформации продуцирующей ткани. Особенно предпочтительным является использование сигнальных пептидов эндоплазматического ретикулума сети или клеточного транспорта. Проведенная по настоящему изобретению работа показала, что вышеуказанная проблема сомаклональных изменений, с которой сталкиваются в клеточных культурах, в фотоавтотрофных жидких культурах мхов не возникает. Кроме того, мхи, используемые в соответствии с изобретением, имеют преимущество над другими системами своей четкой сменой строго определенных стадий дифференцировки (хлоронема, каулонема, почки, гаметофоры), на которые можно влиять добавлением растительных гормонов (индол-3-уксусная кислота стимулирует развитие каулонемы, изопентениладенин стимулирует развитие почек) (см., например, N.W.Ashton et al., Analysis of qametophytic development in the moss, Physcomitrella patens, using auxin and cytokinin resistant mutants, Planta, 144, стр. 427-435 (1979)). Таким образом, в биореакторных культурах возможна направленная экспрессия гетерологичных белков в определенной стадии дифференцировки, и особенно предпочтительным в соответствии с изобретением является синхронное деление чистой и, таким образом, гомогенной культуры хлоронемы по причине контролируемой продукции одинакового белка в биореакторе и удобства использования гормонозависимого промотора или промотора определенной стадии дифференцировки. В дополнение к такой экспрессионной системе также в соответствии с изобретением можно использовать индуцибельную промоторную систему, в частности, для получения белков с коротким периодом полураспада или которые являются цитотоксическими, особенно предпочтительным является использование 1’-промотора Agrobacterium tumefaciens. Культивирование мхов, предложенных в соответствии с изобретением для производства гетерологичных белков в экономических перспективах, может осуществляться, например, с использованием Physcomitrella в объемах с порядками величины от 20 мл до 6-10 л и выше во встряхиваемых культурах или в аэрируемых стеклянных емкостях (см., например, R.Reski, Zell-und molekularbiologische Untersuchungen der cytokinin-induzierbaren Gewebedifferenzierung und Chloroplastenteilung bei Physcomitrella patens (Hedw.) B.S.G., [Cell- and molecularbiological studies of the cytokinin-inducible tissue differentiation and chloroplast division in Physcomitrella patens (Hedw.) B.S.G.], Ph.D. thesis. Университет Гамбурга (1990)). Так как используется культура дифференцированных фотоаутотрофных растений, среда не нуждается ни в добавлении растительных гормонов, ни витаминов, ни cахаров. По сравнению со сложной средой, которая требуется, например, для культур животных клеток, стоимость снижается в 100 раз. В соответствии с изобретением было выяснено, что выход биологически активного гетерологичного белка в культуральной среде может быть увеличен в 35 раз в присутствии ПВП, вот почему в способе по изобретению предпочтительно использование ПВП в культуральной среде. Подробная информация по культивированию в биореакторах других мхов, которые являются подходящими в соответствии с изобретением, таких как, например, Leptobryum pyriforme и Sphagnum magellanlcum, описана в предшествующем уровне техники (см., например, Е.Wilbert, Biotechnologische Studien zur Massenkultur von Moosen unter besonderer Berucksichtigung des Arachidonsaurestoffwechsels [Biotechnological studies concerning the mass culture of mosses with particular consideration of the arachidonic acid metabolism], Ph.D. thesis. Университет Майнца (1991); H. Rudolph и S. Rasmussen, Studies on secondary metabolism of Sphagnum cultivated in bioreactors, Crypt. Bot., 3, стр. 67-73 (1992)). Особенно предпочтительно для целей по настоящему изобретению использование Physcomitrella, в частности, поскольку все стандартные молекулярно-биологические методы созданы для этого организма (см. обзор R. Reski, Development, genetics and molecular biology of mosses, Bot. Acta, 111, стр. 1-15 (1998)). Подходящие трансформирующие системы были разработаны для биотехнологического использования Physcomitrella для получения гетерологичных белков. Например, проводились успешные трансформации посредством прямого переноса ДНК в протонемную ткань, используя бомбардирующую частицами пушку. Также может успешно применяться перенос ДНК в протопласты мха, обусловленный ПЭГ. Этот метод трансформации был неоднократно описан для Physcomitrella и приводит как к временным, так и к стабильным трансформантам (см., например, К.Reutter and R. Reski, Production or a heterologous protein in bioreactor cultures of fully differentiated moss plants, Pl. Tissue culture, @ Biotech., 2, стр. 142-147 (1996)). Хотя настоящее изобретение главным образом подходит для получения любого белкового вещества, получение фармацевтически важного белка здесь демонстрируется на примере человеческого фактора роста эндотелия сосудов (VEGF). VEGF был впервые выделен N.Ferrara и W.J.Henzel (Pituitary follicular cells secrete a novel heparin-binding growth factor specific for vascular endothelial cells, Biochem. Biophys. Res. Commun., 161, стр. 851-858 (1989)) и описан как регуляторный фактор, контролирующий ангиогенез и деление эндотелиальных клеток при нормальных физиологических условиях (N.Ferrara et al., The vascular endothelial growth factor family of polypeptides, J. Cell. Biochem., 47, стр. 211-218 (1991)). Авторы также продемонстрировали, что этот фактор роста обладает высокой специфичностью к эндотелиальным клеткам сосудов и неактивен по отношению к другим типам клеток. VEGF представляет собой гомодимерный гликопротеин, связанный дисульфидными мостиками. Известны четыре различных формы человеческого VEGF. Четыре изоформы имеют протяженность в 121, 165, 189 и 206 аминокислот и образуются альтернативным сплайсингом PHKVEGF. VEGF206 обнаружен только в кДНК печени зародыша, а транскрипты VEGF121 , VEGF165 и VEGF189 обнаружены в большинстве опухолевых клеток и опухолевых тканях. Хотя все изоформы VEGF имеют лидерную последовательность для секреции, эффективно секретируются только две наименьшие формы (см., например, G.Martiny-Baron и D.Marme, VEGF-mediated tumor angiogenesis: A new target for cancer therapy, Curr. Opin. Biotechnol., 6, стр. 675-680 (1995)). До сих пор требуются адекватные количества VEGF как для развития и усовершенствования существующих подходов в терапии опухолей, так и для изучения VEGF. На ранних стадиях работы, проведенной в контексте настоящего изобретения, единственным описанным было рекомбинантное получение VEGF в клетках насекомых посредством бациловирусной экспрессионной системы (например, B.L.Fiebich et al., Synthesis and assembly of functionally active human vascular endothelial growth factor homodimers in insect cells, Eur. J. Biochem., 211, стр. 19-26 (1993)). Saccharomyces cerevisiae (S.Kondo et al., The shortest isoform of human vascular endothelial growth factor/vascular permeability factor (VEGF/VPF121) produced by Saccharomyces cerevisiae promotes both angiogenesis and vascular permeability, Biochim. Biophys. Acta, 1243, стр. 195-202 (1995)), дрожжи Pichia pastoris (D.Mohanraj et al., Expression of biologically active human vascular endothelial growth factor in Yeast, Growth factors, 12, стр. 17-27 (1995)) и Escherichia coli (G. Siemeister et al., Expression of biologically active isoforms of the tumor angiogenesis factor VEGF in Escherichia coli, Biochem. Biophys. Res. Commun., 222, стр. 249-255 (1996)) являются дополнительными продуцирующими организмами. Во всех этих рекомбинантных системах был получен биологически активный VEGF. Однако экспрессионная система Е. coil усложнена в отношении очистки и восстановления белка, так как последний упакован в тельца включения. Примеры Краткое изложение Разработка контролируемых массовых культур Physcomitrella patens (Reutter and Reski, loc. cit.) и способов переноса ДНК в мох Physcomitrella patens (К.Reutter, Expression heterologer Gene in Physcomitrella patens (Hedw.) B.S.G. [Expression of heterologous genes in Physcomitrella patens (Hedw.) B.S.G.], Ph.D. thesis. Университет Гамбурга (1994)) создает основные предпосылки для биотехнологического использования этого растения. Работа, проведенная вначале, показала долговременную стабильность интеграции, используя трансгенные линии Physcomitrella, полученные от Reutter (loc. cit., 1994). Экспрессия гетерологичных генов npt II и gus, которые использовались в качестве примера для этой цели, по прошествии четырех лет до сих пор обнаруживается. Была оптимизирована биореакторная культура Physcomitrella. Была разработана мешалка, которая осуществляла измельчение протонем и, таким образом, обеспечивала требуемую гомогенность культуры при постоянных скоростях 300-500 об/мин. Таким образом, стало возможным стандартизировать взятие образцов. В то же время поступающий воздух распределялся более равномерно в жидкой культуре. При этих условиях было показано, что образование биомассы и белка без регуляции рН извне проходило так же, как при регуляции рН; неожиданно оказалось, что регуляция, следовательно, не нужна. Еженедельную продукцию биомассы, 500 мг сухого веса или 22 мг общего белка на литр, получали в полусерийных условиях. Это означает увеличение производительности биомассы в пять раз по сравнению с обычными культурами в 5-литровых стеклянных колбах. Снижение концентрации соли среды Кноп на одну десятую часть приводило к тем же данным и, таким образом, к снижению стоимости. Добавление 5 мМ тартрата аммония ускоряло развитие биомассы из-за укорочения лаг-периода. Одновременно добавление тартрата аммония давало культуры, которые включали фактически только хлоронемные клетки. Было показано при помощи проточной цитометрии, что фактически сто процентов клеток в этих культурах находились в G2/M фазе клеточного цикла. Этот результат был подтвержден дополнительными физиологическими исследованиями с ауксином и исследованиями с мутантами определенной стадии дифференцировки cal112 и саl113, и были сделаны выводы, что каулонемные клетки большую часть времени находятся в G1/G0 фазе, в то время как хлоронемные клетки находятся преимущественно в G2/M фазе. В качестве возможного индуцибельного промотора мхов рассматривался 1’-промотор агробактерий. В качестве маркерного гена использовался ген Ген 121 аминокислотной сплайсированной формы человеческого фактора роста эндотелия сосудов (VEGF121) трансформировали в Physcomitrella с уровнем трансформации 0,5×10-5 и 3,3х×10-6 . Наконец, ген клонировали за конструктивным промотором 35S и в трансформирующий вектор pRT99, который пригоден для растений. Во втором подходе дополнительно клонировали последовательность, кодирующую соответствующий транспортный белок ER человека. Подвергая полученные стабильные трансформанты Southern-анализу, подтверждали интеграцию гетерологичной ДНК и описывали тип интеграции. Nothern-анализ подтвердил присутствие nptll и двух транскриптов VEGF в этих трансформантах. Экспрессия VEGF121 в клетках мха была показана непрямой иммунофлуоресценцией. Белок однозначно локализовался в клетках, что было показано с помощью конфокального лазерного сканирующего микроскопа. Для трансформантов без транспортного пептида эти исследования показали, что белок локализован, в частности, в цитоплазме. В трансформантах, содержащих транспортный пептид ER, белок может находиться в области ядра и в апикальных областях апикальных клеток, областях с очень высоким содержанием ER. Биологическую активность гетерологичного белка, полученного в соответствии с изобретением, проверяли с помощью анализа ELISA и двух анализов на функциональную активность, используя белок VEGF, полученный из питательной среды. Материалы и методы: Если иначе не указано в тексте, используемые химические соединения были чистые для анализа и их получали от Fluka (Neu-Ulm), Merck (Darmstadt), Roth (Karlsruhe), Serva (Heidelberg) и Sigma (Deisenhofen). Растворы готовили на очищенной, апирогенной воде, здесь обозначенной как Н2О, из системы водной очистки Milli-Q (Millipore, Eschborn). Рестрикционные эндонуклеазы, ДНК-модифицирующие ферменты и биологические наборы получали от AGS (Heidelberg), Amersham (Braunschweig), Applied Biosystems (Weiterstadt), Biometra (Gottingen), Boehringer Mannheim GmbH (Mannheim), Genomed (Bad Oeynhausen), New England Biolabs (Schwalbach/Taunus), Novagen (Madison, Wisconsin, USA), Pharmacia (Freiburg), Qiagen (Hilden) и Stratagene (Heidelberg). Если не определено иначе, их использовали в соответствии с инструкциями изготовителя. Векторы и конструкции Плазмида pCYTEXP-VEGF121 является производной плазмиды pCYTEXPl (T.N. Belev et al., A fully modular vector system for the optimization of gene expression in Escherichia coil, Plasmid, 26, стр. 147-150 (1991)), в которую интегрировали кДНК человеческого VEGF121 для экспрессии в Е. coil. Используя рестрикционные эндонуклеазы Nde I и Sal I из pCYTEXP-VEGF 121, вырезали кДНК VEGF121, очищали, делали ей тупые концы и клонировали в рестрикционный сайт Sma I pRT101 (R. Topfer et al., A set of plant expression vectors for transcriptional and translational fusions, Nucleic Acids Res., 15, стр. 5890 (1987)) между промотором 35S и полиаденилированной последовательностью CaMV. Полученную таким образом кассету снова разрезали Hin dIII и клонировали в сайт рестрикции Hin dIII трансформирующего вектора pRT99. pRT99 содержит не только полилинкерный сайт рестрикции, но также ген фосфотрансферазы неомицина под регуляцией промотора 35S и соответствующую полиаденилированную последовательность CaMV (R. Topfer et al., Versatile cloning vectors for transient gene expression and direct gene transfer in plant cells, Nucleic Acids Res., 16, стр. 8725 (1988)). Этот ген дает устойчивость к антибиотику G418 стабильно трансформированным растениям. Плазмиды реплицировались в штамме DH5 Используя рестрикционные ферменты Ваm HI и Вgl II, кДНК вырезали из вектора pVE-121, который был первоначально создан для экспрессии VEGF121 в клетках насекомых и который кроме последовательности VEGF121 включает ДНК, кодирующую природный транспортный пептид, который, в свою очередь, в системе животных клеток опосредует секрецию в среду из эндоплазматического ретикулума (Fiebich et al., Synthesis and assembly of functionally active human vascular endothelial growth factor homodimers in insect cells, Eur. J. Biochem., 211, стр. 19-26 (1993)) и, используя pRTlOl, клонировали в pRT99 и проверяли. Плазмида pNA201 является производным бинарного вектора pBI101 (A.R.Jefferson et al., Assaying chimeric genes in plants: The GUS gene fusion system. Plant Mol. Rep., 5, стр. 387-405 (1987)). В качестве селективного маркера растений она содержит ген nptII под нопалинсинтазным промотором. Ген gus, который также присутствует, регулируется 1’-промотором Agrobacterium tumefaciens. pNA201 удобен для прямой трансформации Physcomitrella patens. Антитела Использовали два различных антитела против белка VEGF. Первым антителом было кроличье анти-VEGF антитело, направленное против синтетического пептида, соответствующего 1-20 аминокислотам нативного человеческого VEGF (Fiebich et al., loc. cit. (1993)). Вторым антителом было моноклональное мышиное антитело против человеческого белка VEGF 121 (R&D Systems, Wiesbaden). Растительный субстрат Использовался штамм дикого типа мха Physcomitrella patens (Hedw.) B.S.G., который был взят из коллекции Genetics Group при Department of General Botany, Университет Гамбурга. Его источником является штамм 16/14, который находится в коллекции H.L.K. Whitehouse в Gransden Wood, Huntingdonshire (Англия) и был субклонирован из споры. Штамм дикого типа выращивали либо в виде жидкой культуры со средой Knop (R.Reski и W.O.Abel, Induction of budding on chloronemata and caulonemata of the moss, Physcomitrella patens, using isopentenyladenine, Planta, 165, стр. 354-358 (1985)), или на твердой среде с 1% агаром Oxoid (Unipath, Basingstoke, Англия). Жидкие среды получали как описано у Reski (loc. cit., 1990). Биореакторная культура Для массовой культивации растительный материал помещали в 7-л круглодонные стеклянные колбы биореактора, оснащенного двойной обшивкой (Applikon Biotek, Knullwald). В системе биореактора сверху через отверстие в крышке в объем культуры вводили конденсатор с воздушным охлаждением для отвода газа, аэрационную трубку, рН-электрод (Conducta, Gerlingen), температурный датчик, устройство для перемешивания, трубку для взятия проб и входа для кислоты, основания и среды. Для культур устанавливали температуру 25°С и контролировали ее посредством соответственно отрегулированной водяной бани, которая присоединялась к двойной обшивке. В эксперименте, который проводили с регулированием рН, рН поддерживали постоянным при 5,8 с помощью единицы титрования. Температурные измерения и регулирование рН проводили с помощью Biocontroller ADI 1030 (Applikon Biotek, Knullwald). Скорость мешалки можно изменять регулятором частоты вращения ADI 1012 (Applikon Biotek, Knullwald). Питательную среду постоянно аэрировали 1 баром воздуха через пористый вдувающий элемент. Для обеспечения стерильности в колбе с культурой все подводящие и отводящие трубки были снабжены устройством для стерилизации фильтрацией (Midisart, 0,2 мкм; Sartorius, Gottingen). Биореакторную культуру помещали в камеру с контролем климата и боковым освещением (белый свет; Osram L 40 W/20; максимально 100 мкмоль-1м-2). Культуру инокулировали 0,5-1 г сырого растительного субстрата на литр культуры биореактора в стерильных условиях. Трубка для взятия образцов находилась на одном уровне с мешалкой, таким образом, гарантируя единообразие отбора образцов во время перемешивания. Небольшие по количеству образцы (<100 мл) обирали стерильным шприцем через соединения типа Luer-lock, в то время как большие образцы отбирали с помощью, например, перистальтического насоса типа 302/3А, оснащенного головкой 501 RI (Sartorius, Gottingen). Хлоронемную культуру дикого типа получали при добавлении в среду Кnор 5 мМ тартрата аммония. Выход биологически активного секретируемого гетерологичного белка может быть существенно повышен в присутствии в питательной среде стабилизатора поливинилпирролидона (ПВП). Дифференцировка каулонемы, которая начинается в процессе развития протонемы, может быть индуцирована и усилена добавлением извне физиологических количеств ауксина в подходящих концентрациях, составляющих, например, 5 мкмоль/л индол-3-уксусной кислоты (ИУК). Для обеспечения давления отбора в жидкой культуре трансформантов добавляли 50 мг/л антибиотика G418 (Calbiochem, Bad Soden) в среду Кnор. Наконец, непосредственно перед измельчением каждые десять дней культуру удаляли фильтрацией через стерильный 100 мкм решетчатый фильтр (Wilson Sieves, Nottingham, Англия) и переносили в колбы Эрленмейера, наполненные селективной средой. Для экспериментов с получением питательных элементов среду Кnор разводили 1:10 водой. Трансформация Выбранный способ трансформации представляет собой ПЭГ-опосредованный прямой перенос ДНК в протопласты, как описано у Reutter и Reski (loc. cit., 1996). Для каждой трансформации на 3×105 протопластов использовали 50 мкг плазмидной ДНК. Обновление протопласта и отбор стабильных трансформантов проводили, если не указано иного, как описано у Reutter и Reski (loc. cit., 1996). Непрямая иммунофлуоресценция Буфер: MSB: 100 мМ PIPES; 5-10 мМ EGTA, 5 мМ MgSO4, pH 6,8 F-MSB: MSB + 5% ДМСО E-MSB: MSB + 5% ДМСО + 5% Nonidet W-MSB: MSB разбавленный 1:2 Н2О (промывочный буфер) Ферментный раствор: 1% целлюлозы, 1% пектиназы, 2% дризелазы в MSB, pH 5,6 (все от Sigma, Deisenhofen) Протонему мха фиксировали в 1,25% глутаровом альдегиде с F-MSB (об/об), инкубировали более 10 минут и быстро промывали W-MSB. Затем протонему инкубировали 40 минут с 2% параформальдегидом в MSB (об/об) и 3 раза промывали W-MSB (1 раз полоскали; 2 раза промывали по 5 минут). Свободные альдегиды, которые могут быть не удалены промывкой, удаляли добавлением MSB и твердого гидрата бора на кончике шпателя и инкубировали в течение 10 минут. Три раза промывали W-MSB для удаления гидрида бора. На следующей стадии для проницаемости клеточных стенок добавляли ферментный раствор и инкубировали 10 минут. Ферментную реакцию прерывали изменением рН (MSB, рН 6,8). Смесь снова промывали 3 раза W-MSB. Хлорофиллы экстрагировали инкубацией с раствором детергента в течение 120 минут. Раствор затем удаляли, промывая три раза W-MSB. После такой подготовки протонему мха можно инкубировать с первым антителом (анти-VEGF; разбавление 1/50). Инкубировали при 37°С в течение 45 минут. После трех промывок W-MSB добавляли меченые вторые антитела (антикроличьи или антимышиные; разведение 1/30; меченные флуоресцентным изотиоцианатом (ФИТЦ) (Molecular Probes, Leiden, Netherlands)) и инкубировали в течение 45 минут при 37°С. Дополнительно к стадии 3-х промывок, описанной выше, смесь один раз промывали W-MSB + 0,1% Triton. Протонемы затем переносили в W-MSB и хранили при 4°С, по крайней мере, в течение ночи. Вещество оценивали с помощью конфокального лазерного сканирующего микроскопа (CLSM) типа TCS 4D (Leica Lasertechnik, Heidelberg) и программного обеспечения Scanware 5,0 (Leica Lasertechnik, Heidelberg). Для анализа образцов с помощью CLSM их помещали на предметное стекло в закрепляющий раствор (Dabco, Sigma, Deisenhofen). Возбуждение флуоресцентного красителя ФИТЦ, присоединенного ко второму антителу, осуществляли аргон-криптоновым лазером при длине волны 488 нм. ФИТЦ испускает свет при длине волны 528 нм. Анализ ELISA VEGF белок в культуральной среде, который образуется в соответственно трансформированных растениях мха, качественно и количественно определяли обычными методами, используя анализ ELISA и вышеуказанные антитела. В анализе ELISA непосредственно использовали 200 мкл культуральной среды. Анализы функциональной активности Биологическое действие рекомбинантного VEGF, полученного из культуральной среды, проверяли в митогенетическом анализе (Miyazono et al., Purification and properties of an endothelial cell growth factor from human platelets, J. Biol. Chem., 262, стр. 4098-4103 (1987)) и в “анализе ангиогенеза 13-дневной хорион-аллантоидной мембраны” (Wilting et al., A morphological study of the rabbit corneal assay, Anat. Embryol., 183, стр. 1167-1174 (1991)). Культуральную среду заранее подвергали ультрафильтрации, затем лиофилизовали и впоследствии ресуспендировали в буфере. Если желательно, в качестве дополнительной стадии очистки можно использовать катионную колонку. Индукция 1’-промотора Способность к индукции ауксином 1’-промотора анализировали 5 мкмоль/л индол-3-уксусной кислоты (ИУК). Через пять дней после трансформации 100 мкл протопластных аликвот из реакции трансформации с pNA201 переносили в 96-луночный микротитровальный планшет (Nunc, Wiesbaden). Протопластные суспензии культивировали в течение пяти часов с ИУК (конечная концентрация = 5 мкМ). Эксперименты с индукцией оценивали непосредственно после периода инкубации качественным анализом Качественный анализ В качественном анализе определяли активность Субстратный буфер: 50 мМ К3Fе(СN)6 50 мМ Na2HPO4 50 мМ K4Fe(CN) 6 1% (об/об) Triton X-100 50 мМ NaH2 PO4 10 мМ EDTA, pH 7,0 4 мг/мл ПВП (М.м. 10000) Окрашивающий раствор: 12,5 мг 5-бром-4-хлор-3-индолил-глюкуроновой кислоты (Biomol, Гамбург) растворяли в 250 мкл N,N-диметилформамид/50 мл субстратного буфера. Протонемы мха и протопласты мха в среде Кnор или восстанавливающей среде инкубировали до 72 часов при 37°С с равным объемом окрашивающего раствора и после этого немедленно оценивали под микроскопом. Результаты Гомогенность биореакторной культуры; взятие образцов Только гомогенные культуры обеспечивают стандартизацию отбора образцов из клеточной культуры. После продолжительного культивирования рост протонемы в длинных клеточных филаментах часто приводит к агрегации клеток и, таким образом, к негомогенному распределению растительного субстрата в жидких культурах. Чтобы избежать такой агрегации, протонемы измельчают через определенные промежутки времени - в биореакторе через день, начиная с 10 дня, а во встряхиваемой культуре через каждые 12 дней, используя высокоскоростные мешалки/гомогенизаторы. Для осуществления постоянных условий в биореакторе, наряду со стандартизацией отбора образцов даже после длительного периода культивации, рекомендуется модифицировать трехлопастную турбинную мешалку путем заточки граней лопастей мешалки, таким образом преобразуя их в ножевую поверхность. Постоянное “перемешивание” при 300-500 об/мин, таким образом, делает возможным работу с гомогенными биореакторными культурами. Развитие биомассы (в DW [мг/л]) свыше 35 дней (840 часов) при 500 об/мин в контрольных культурах с турбинной мешалкой и в биореакторных культурах, перемешиваемых мешалкой с ножевой поверхностью, одинаковое. В каждом случае гомогенность культуры оценивалась сравнением шести параллельных образцов. Сухой вес растительного субстрата из образцов объемом 100 мл определяли как параметр сравнения. При использовании немодифицированной турбинной мешалки среднеквадратичное отклонение увеличивалось при увеличении концентрации биомассы. Как следствие перемешивания мешалкой с ножевой поверхностью, среднеквадратичное отклонение взятых образцов оставалось низким. Это позволяет сделать вывод, что единообразная гомогенная культура, растущая больше 35 дней, может быть получена при использовании модифицированной мешалки. Исследование способности к индукции 1’-промотора В плазмиде pNA201, 1’-промотор находится перед геном gus и действует как регулирующий элемент. Известный анализ Реакции трансформации подвергали анализу Получение трансформирующих векторов для продукции VEGF В Physcomitrella для трансформации кДНК VEGF121 без лидерной последовательности, обозначенной здесь и далее как VEGFC, и кДНК VEGF121 с лидерной последовательностью, обозначенной здесь и далее как VEGFP, необходимо клонировать последовательности между промотор/терминаторной единицей, подходящей для растений. Для этой цели выбраны промотор 35S CaMV и соответствующий сигнал полиаденилирования. Соответственным образом полученная последовательность kDNAVEGF клонируется в рестрикционный сайт Sma I полилинкерного сайта рестрикции вектора pRT101. Полученные векторы (pRT101VEGFC 3 и VEGFP 21) секвенировали с праймером, полученным из терминальной области промотора 35S, и проверяли правильность интеграции между промотором и сайтом полиаденилирования. Полученные кассеты вырезали ферментом рестрикции Hin dIII и клонировали в функциональный трансформирующий вектор pRT99 в Hin dIII сайт (pRT99VEGFC 3 и VEGFP 21). Ориентация кассет относительно NPTII кассеты может быть определена рестрикцией Sma I и Hinc II. В случае ориентации промотор-сигнал полиаденилирования получали фрагменты 5250 (VEGFC) и 5380 п.н. (VEGFP), в то время как в случае обратной ориентации получали фрагменты 1100 (VEGFC)/1230 (VEGFP) и 4150 п.н. (VEGFC и Р). Рестрикционный анализ показал только 5250/5380 п.н. фрагмент, и, то есть, VEGFC/P кассеты относительно гена nptll pRT99 были встроены с ориентацией промотор-сигнал полиаденилирования. Трансформация VEGFC в Physcomitrella После неоднократной замены среды Кпор селекционной средой абсолютный уровень трансформации для трансформации конструкции VEGFC в протопласты дикого типа с постоянными стабильными трансформантами равнялся 0,5×10-5. Демонстрация интеграции трансформирующей плазмиды Успешная интеграция в геном растения демонстрировалась Southern-гибридизацией. Использовали зонды, во-первых, фрагмента Nco I гена npt II из pRT99 и, во-вторых, фрагмента Nde I/Sal I VEGFC pCYTEXP-VEGF121. Для подтверждения успешной интеграции плазмидной ДНК в геном растения определяли сигналы во всей неразрезанной ДНК трансформантов. Любую из всех полученных кассет 35S VEGFC PolyA, даже после интеграции, тестировали рестрикцией Нin dIII. Если кассета осталась неповрежденной при интеграции, этот фермент вырезал из всей ДНК фрагмент с 1100 п.н. Для всех трансформантов профиль гибридизации с зондом VEGFC обнаруживал фрагмент с 1100 п.н. Таким образом, была показана интеграция полной единицы экспрессии VEGFC, необходимой для правильной транскрипции и экспрессии VEGF121 во мхе. Демонстрация транскрипции гетерологичных генов Транскрипты гетерологичных генов VEGFC и NPTII трансформантов обнаруживали нерадиоактивной системой детекции РОЕТ, используя зонды VEGF и NPT II. Обнаруженные на флуорограмме размеры транскриптов, 760 нуклеотидов для VEGFC транскрипта и 1100 нуклеотидов для NPT II транскрипта, находятся в пределах порядка величины, ожидаемой для каждого случая. Как и ожидалось, ни один из двух гетерологичных транскриптов не был обнаружен в WT контроле. Анализ трансформантов с человеческим транспортным пепгидом При ПЭГ-опосредованном переносе ДНК, 50 мкг pRT99P 21 плазмидная ДНК на реакцию трансформации, получали постоянно стабильные на селекционной среде трансформанты. Уровень стабильных трансформантов составлял 3,3×10-6. Демонстрация интеграции трансформирующей плазмиды Интеграция вышеописанных трансформантов с транспортным белком демонстрируется с помощью Southern-гибридизации, как описано выше, используя описанные зонды, а гибридизацией всей ДНК, разрезанной Hin dIII, с зондом VEGF обнаруживается фрагмент 1230 п.н.: демонстрация полной интеграции кассеты 35S VEGFP PolyA. Демонстрация транскрипции гетерологичных генов Как NPTII, так и VEGFP транскрипты могут быть обнаружены методом, описанным выше. Обнаружение человеческого VEGF121 в трансгенных клетках мха с использованием конфокального лазерного сканирующего микроскопа В этом методе тестируемый белок был мечен непосредственно в закрепленных клетках. Оценку проводили под конфокальным лазерным сканирующим микроскопом, который имеет улучшенную разрешающую способность по сравнению с обычным оптическим микроскопом. Рекомбинантный белок VEGF121, если он обнаруживается в клетках мха, должен скапливаться в цитоплазме VEGFC трансформантов. Если транспортный пептид во мхе работает как сигнальный, то в трансформантах VEGFP он должен обнаруживаться в ER системе. Экспрессия человеческого VEGF121 в трансгенных клетках мха была успешно продемонстрирована методом непрямой иммунофлуоресценции и оценкой, выполняемой с помощью компьютера, конфокальным лазерным сканирующим микроскопом. Кроме того, было показано, что в трансгенном мхе в присутствии соответствующего человеческого транспортного пептида VEGF121 успешно транспортируется в эндоплазматический ретикулюм. Анализ на присутствие VEGF в культуральной среде С помощью ELISA анализировали 200 мкл аликвоту культуральной среды с добавлением ПВП и показали, что растения мха, трансформированные экспрессионной кассетой, включающей последовательность, кодирующую транспортный пептид, способны к высвобождению VEGF в среду. Положительные результаты позволили сделать вывод о присутствии функционального белка VEGF. Анализ биологической активности Оба анализа, используемые для проверки биологической активности VEGF белка, высвобождаемого в культуральную среду, дают положительные результаты и подтверждают, что VEGF, полученный в соответствии с изобретением, может быть получен из питательной среды с желаемым биологическим действием. Формула изобретения1. Способ получения гетерологичных белковых веществ из растительного субстрата, отличающийся тем, что в качестве растительного субстрата используют протонему ткани мха и продуцируемые белковые вещества выделяют из культуральной среды без повреждения продуцирующих тканей или клеток. 2. Способ по п.1, отличающийся тем, что белковые вещества, высвобождающиеся в культуральную среду, являются биологически активными. 3. Способ по п.1 или 2, отличающийся тем, что используемая культуральная среда не содержит сахаров, витаминов и фитогормонов или их функциональных фрагментов. 4. Способ по любому из пп.1-3, отличающийся тем, что ткань мха выбрана из группы, состоящей из мхов и печеночников. 5. Способ по п.4, отличающийся тем, что ткань мха выбрана из группы, состоящей из Physcomitrella, Funaria, Sphagnum и Ceratodon. 6. Способ по п. 4, отличающийся тем, что ткань мха выбрана из группы печеночников, состоящей из Marchantia и Sphaerocarpos. MM4A Досрочное прекращение действия патента из-за неуплаты в установленный срок пошлины за поддержание патента в силе Дата прекращения действия патента: 28.09.2012 Дата публикации: 20.07.2013 Популярные патенты: 2056755 Способ регулирования роста овощных культур ... растений томатов препаратом Сапфир приводит к более чем двухкратному снижению содержания нитратов в плодах. П р и м е р 4. 1 кг отходов производства пива, содержащих сухую массу корешков проростков ячменя, экстрагировали 10 л 70%-ного этилового спирта в течение 24 ч при 4оС. Экстракт упаривали под вакуумом до водного остатка. Водный остаток дважды промывали равными объемами этилового эфира, доводили рН до 7,8 с помощью КОН и экстрагировали равным объемом н-бутанола, после чего удалили н-бутанол под вакуумом до исчезновения запаха органического растворителя. Анализ полученного таким образом препарата (препарат Сапфир) методом спектроденситометрической тонкослойной хроматографии ... 2025945 Способ выращивания насаждений сосны ... обеспечивающую формирование древостоя согласно предложенному способу. П р и м е р 4. В 1946 г. в Балаклейском лесхоззаге Б.И.Гавриловым заложен опыт в 8-летнем чистом, абсолютно одновозрастном молодняке на площади 1,9 га, созданных с размещением посадочных мест 1,5х0,75 м (10 тыс. шт/га). При закладке опыта в культурах сосны был вырублен каждый второй ряд и произведено разреживание оставшихся рядов, а в последствии - обрезка сучьев на деревьях до высоты 4 м. Контролем служило насаждение с производственным режимом выращивания с естественным очищением стволов от сучьев, созданное по аналогичной схеме посадки в идентичных условиях местопроизрастания - свежей субори - В2. ... 2422377 Биоцидный концентрат ... мкм, а затем динамически закалены в результате падения в воду. Динамический контроль межэлектродного промежутка осуществляется при помощи электропривода, содержащего микрометрический винт и связанного с датчиком. Технический результат заключается в улучшении служебных характеристик биоцидного концентрата, а также расширении технологических возможностей по его применению. Изобретение относится к технологии и средствам обработки воды, а более конкретно обеззараживанию питьевой воды посредством насыщения наночастицами биоцидного действия, полученными при импульсных дуговых разрядах в обрабатываемой воде. Уровень данной области техники характеризует консервант, описанный в патенте ... 2112361 Контроллер программируемого управления поливом ... 2 выход 10, 22, 23, вариант 2) на 2 участок будет вылито 1000 л воды, первый - 3000 л, затем вновь на второй 4000 л, первый 2000 л и т.д. При положении переключателя 12 влево (как изображено на фиг. 1) выходы схем И 8 и 9 будут подключены к схеме И 15. Схемы 8 и 9 открываются в противофазе от триггера 23. Схема 8 пропускает импульсы от технологического датчика, а схема 9 - от таймера. Период от начала пуска до подачи команды (фиг. 2, поз. 9, вариант 3) характеризует реальное время до начала пуска поливной машины, начала технологического процесса полива, от первой до второй команды (фиг. 2, поз. 8, 22 вар. 3) - включение поливной машины (полив с заданным количеством осадков, ... 2126616 Устройство управления навесной системой трактора ... договор: 02.02.2011 РД0076005 Изменения: Срок действия продлен до 31.12.2012 г. Дата публикации: 10.03.2011 QZ4A Внесение изменений в зарегистрированный договор Дата и номер государственной регистрации договора, в который внесены изменения: 20.07.2005 № РД0001102 Вид договора: лицензионный Лицо(а), передающее(ие) исключительное право: Республиканское унитарное предприятие "Минский тракторный завод" (BY) Лицо, которому предоставлено право использования: Открытое акционерное общество "Производственное объединение Елабужский автомобильный завод" (RU) Дата и номер государственной регистрации изменений, внесенных в зарегистрированный договор: ... |
Еще из этого раздела: 2021671 Машина для уборки льна-долгунца 2303347 Способ ведения виноградных кустов 2485762 Ракета для активного воздействия на облака 2010519 Способ биологической борьбы с вредителями растений 2167648 Средство для защиты от укусов кровососущих насекомых (варианты) и способ его получения 2440708 Комбинированное устройство для ротационного внутрипочвенного рыхления 2028763 Измельчитель древесной поросли 2235450 Малогабаритная машина для обескрыливания, очистки и сортирования лесных семян 2264065 Способ возделывания сельскохозяйственных культур на корм 2502259 Способ получения водорастворимого бактерицидного препарата |
Изобретения в сельском хозяйстве
Обработка почвы в сельском и лесном хозяйствах
Посадка, посев, удобрение
Уборка урожая, жатва
Обработка и хранение продуктов полеводства и садоводства
Садоводство, разведение овощей, цветов, риса, фруктов, винограда, лесное хозяйство
Новые виды растений или способы их выращивания
Производство молочных продуктов
Животноводство, разведение и содержание птицы, рыбы, насекомых, рыбоводство, рыболовство
Поимка, отлов или отпугивание животных
Консервирование туш животных, или растений или их частей
Биоцидная, репеллентная, аттрактантная или регулирующая рост растений активность химических соединений или препаратов
Хлебопекарные печи, машины и прочее оборудование для хлебопечения
Машины или оборудование для приготовления или обработки теста
Обработка муки или теста для выпечки, способы выпечки, мучные изделия
|
|
||
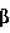 -глюкоронидазы (gus). Во временно трансформированных протопластах мха (уровень трансформации = 3×10-4), после индукции. 5 мкМ индол-3-уксусной кислотой наблюдалась экспрессия гена gus. Ни в каком контроле экспрессии не наблюдалось.
-глюкоронидазы (gus). Во временно трансформированных протопластах мха (уровень трансформации = 3×10-4), после индукции. 5 мкМ индол-3-уксусной кислотой наблюдалась экспрессия гена gus. Ни в каком контроле экспрессии не наблюдалось.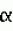 Escherichia coil (J. Sambrook et al., Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory Press, Нью-Йорк (1989)).
Escherichia coil (J. Sambrook et al., Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory Press, Нью-Йорк (1989)).