Способ диагностирования состояний пчелиной семьиПатент на изобретение №: 2161883 Автор: Дрейзин В.Э., Рыбочкин А.Ф., Захаров И.С. Патентообладатель: Курский государственный технический университет Дата публикации: 20 Января, 2001 Начало действия патента: 13 Июля, 1998 Адрес для переписки: 305040, г.Курск, ул. 50 лет Октября 94, КГТУ, ОИС Изображения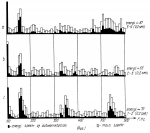   Изобретение предназначено для использования в пчеловодстве и состоит в том, что по издаваемому акустическому шуму пчелиной семьи вычисляют энергетический спектр реализации дискретизованного сигнала, состоящий в том, что исходный аналоговый сигнал предварительно подвергают частотной фильтрации с выделением наиболее информативного частотного диапазона, например 100-600 Гц, затем сигнал дискретизируют и из него выбирают 3-5 коротких реализаций длительностью 0,5-1,0 с, отвечающих условию незначимых различий их дисперсий, и по одной из них рассчитывают энергетический спектр, который делят на 10-12 частотных полос, для которых рассчитывают усредненные значения спектральной плотности и производят распознавание состояния пчелиной семьи путем сопоставительного анализа, усредненных значений спектральной плотности этих же полос частот для диагностируемого состояния пчелиной семьи. 3 ил. , , ОПИСАНИЕ ИЗОБРЕТЕНИЯ К ПАТЕНТУИзобретение относится к области пчеловодства и может быть использовано для диагностирования состояний пчелиной семьи. Известен способ распознавания состояний пчелиной семьи и других коллективных насекомых по издаваемому акустическому шуму путем его спектрального анализа и измерения амплитуд отдельных спектральных составляющих [1, 2]. Его основным недостатком является весьма существенные вариации результатов измерений отдельных спектральных составляющих, полученные по различным реализациям акустического сигнала. В частности, для получения приемлемой достоверности распознавания состояния пчелиной семьи по амплитудному спектру необходимо проводить усреднение измеряемых значений отдельных спектральных составляющих путем интегрирования в течение 30 с (длительность реализации звукового сигнала пчелиной семьи должна быть не меньше 30 с.). Для обработки такой реализации путем спектрального разложения (прямое преобразование Фурье) [3] потребуется значительное время для вычисления. Именно это обстоятельство препятствует использованию числового метода спектрального разложения предварительно дискретизированного акустического сигнала, что могло бы существенно повысить быстродействие и точность измерения спектральных составляющих сигнала. Акустический сигнал пчелиной семьи не является детерменированным [1]. Его можно считать стационарным эргодическим случайным сигналом [4]. Однако для стационарного и эргодического случайного процесса амплитуды его спектрального разложения также являются случайными величинами с определенным законом распределения. Поэтому для практического использования этого способа распознавания состояний пчелиной семьи приходится находить математические ожидания амплитуд отдельных спектральных составляющих, для чего приходится усреднять спектры, полученные по многим реализациям сигнала. Кроме того, в описываемом методе распознавание тех или иных состояний пчелиной семьи предлагалось вести по наличию (или отсутствию максимумов) спектральной плотности в определенных частотных диапазонах. Однако, как видно из представленных в [1] амплитудных спектров, измеренных при различных состояниях пчелиной семьи, такой способ распознавания не будет обладать высокой достоверностью, поскольку а) спектры сигналов, соответствующих конкретным состояниям пчелиной семьи, могут иметь не один, а несколько максимумов спектральной плотности; б) спектры сигналов, соответствующих различным состояниям пчелиной семьи, могут иметь локальные максимумы спектральной плотности в одних и тех же частотных полосах, только для одного состояния они будут глобальными, а для другого - локальными; в) на относительную величину локальных максимумов спектральной плотности и их положение (на оси частот) существенно влияют время суток, календарное время, погодные условия, численность пчелиной семьи, внешние раздражители (шум, тряска, удар и т.д.) и другие факторы, соответствующие моменту измерения спектра акустического сигнала. Поэтому для достоверного распознавания конкретных состояний пчелиной семьи необходимо не просто определить наличие максимума спектральной плотности в данной узкой полосе частот, а провести сопоставительный анализ значений спектральной плотности для нескольких наиболее характерных для распознавания данных состояний областей частот. Величина средней мощности сигнала звуков пчел в элементарном диапазоне частот есть результат усреднения ее по множеству реализаций, и поэтому не является случайной, а определяется функцией спектральной плотности мощности, т. е. энергетическим спектром. Поэтому акустические сигналы пчелиной семьи корректно характеризовать не спектральной функцией его реализации, которая должна иметь длительность не меньше 30 с, а спектральной плотностью мощности, которую можно вычислить из автокорреляционной функции, определяемой по одной очень короткой реализации (значительно меньше 30 с) [4]. Технической задачей, на решение которой направлено предлагаемое изобретение, является обеспечение возможности достоверного распознавания состояния пчелиной семьи по одной короткой реализации акустического сигнала, создаваемого пчелиной семьей. Поэтому первой задачей, которую необходимо решить, является определение минимальной длительности реализаций, по которой можно достоверно вычислить энергетический спектр. Для этого был проведен анализ влияния длительности реализаций на вычисленный по ней энергетический спектр. Результаты этого анализа можно проиллюстрировать на примере. Пример Нами были проведены три эксперимента по выбору минимальной длительности реализаций, осуществлялся анализ трех реализаций одинаковой длительности (фиг. 1 - фиг. 3), взятых из одного и того же сигнала, соответствующего случаю, когда в пчелином гнезде находятся две матки (при создании новой пчелиной семьи матка оказалась на вынутой рамке, с применением этой рамки создавался отводок, матка не была замечена пчеловодом и в новую пчелиную семью была подсажена в клетке другая матка, в итоге новая матка не принималась). Получены реализации, которые расположены друг за другом. Из сопоставления полученных спектров сделаны следующие выводы: 1. При длительности реализаций, равной 0,2 с, амплитудные и энергетические спектры соседних реализаций существенно отличаются, причем энергетические спектры коррелируют с амплитудными (фиг. 1). 2. При последовательном увеличении длительности реализаций до 0,5 с и 1 с различия между энергетическими спектрами уменьшаются. 3. В энергетических спектрах реализаций длительностью в 1 с основная мощность сигнала локализуется в узком частотном диапазоне (140-160 Гц), в то время как амплитудные спектры хотя и имеют максимум на этих частотах, но распределены более равномерно по всему анализируемому частотному диапазону (от 100 до 600 Гц). Однако сигналы общего шума пчел могут содержать звуки отдельных пчел (если они издают звуки в непосредственной близости от микрофона), существенно искажающие спектр сигналов. Поскольку для уменьшения времени вычислений необходимо использовать как можно более короткие реализации (порядка 1 с), то необходим выбор реализаций, свободных от доминирующего звука отдельных пчел. Отбор таких реализаций представляет вторую задачу, которую необходимо решить. Критерием отбора пригодных реализаций является идентичность их моментов распределения. Использовать для этих целей моменты первого порядка (математические ожидания) нельзя, т.к. сами сигналы являются центрированными (т.е. их математические ожидания равны нулю). Из остальных моментов наиболее просто вычисляются моменты второго порядка (т.е. дисперсия). Он и принят в качестве критерия идентичности реализаций. Практически проводится дискретизация и запись 3-5 последовательных реализаций длительностью в 1 с и вычисляются их дисперсии. Если дисперсии отдельных реализаций отличаются незначимо, то по любой из них находится автокорреляционная функция, затем проводится ее спектральное разложение, и по соотношению отдельных составляющих этого спектрального разложения судят о состоянии пчелиной семьи. Согласно [3, 4] спектральное разложение (прямое преобразование Фурье) от автокорреляционной функции имеет вид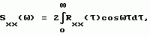 (1) где (1) где 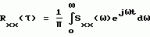 (2) есть автокорреляционная функция случайного сигнала и представляет собой функцию спектральной плотности мощности случайного сигнала. Для стационарных эргодических процессов эта функция не случайна и остается постоянной для любой реализации случайного сигнала. Значения отдельных спектральных составляющих этой функции, вычисленные по любой из реализаций одного и того же случайного сигнала, имеют гораздо меньшие вариации, чем амплитуды тех же спектральных составляющих исходного сигнала. Наиболее приемлемым способом определения функции спектральной плотности мощности является числовой, поскольку аналоговыми методами получить автокорреляционную функцию случайного сигнала сложно. Однако при численном методе определения энергетического спектра случайного сигнала для очень коротких реализаций сигнала существенное значение имеют частота дискретизации исходного аналогового сигнала, величина шага квантования при расчете автокорреляционной функции и частотное разрешение при расчете по ней энергетического спектра [4]. При выборе этих величин необходимо руководствоваться необходимостью обеспечения достаточной достоверности получаемого спектра, т.е. минимальных различий спектров, получаемых по коротким реализациям одного и того же стационарного случайного процесса, и минимум затрат машинного времени, необходимого для анализа спектра. Выбор частоты дискретизации исходного сигнала обычно осуществляют в соответствии с теоремой Котельникова. Однако поскольку наиболее информативной частью спектра акустических шумов пчелиной семьи является диапазон от 100 до 600 Гц, в то время как частотный диапазон самих сигналов простирается до десятков кГц, то выбор частоты дискретизации здесь не является тривиальным вопросом. Если выбрать частоту дискретизации, ориентируясь на верхнюю границу интересующего нас частотного диапазона (fд = 1200-1500 Гц), то наличие в сигнале частот свыше 600 Гц приведет к дополнительному разбросу спектров отдельных реализаций одного и того же сигнала. С другой стороны, увеличение частоты дискретизации будет приводить к резкому увеличению объема вычислительных работ и необходимых ресурсов оперативной памяти ЭВМ, что нежелательно. Поэтому наилучшим решением является сохранение низкой частоты дискретизации (1200-1500 Гц) при предварительной частотной фильтрации исходного аналогового сигнала с выделением из него интересующего нас частотного диапазона (100-600 Гц). При этом дополнительного разброса спектров отдельных реализаций происходить не будет. Шаг квантования при расчете автокорреляционной функции обычно выбирается равным шагу дискретизации исходного сигнала. Шаг квантования по частоте при расчете энергетического спектра должен выбираться из следующих соображений. Для удобства анализа и надежного распознавания отдельных состояний пчелиной семьи по энергетическому спектру желательно разделить весь частотный диапазон на 10-12 полос и получать усредненные значения спектральной плотности для этих полос. В этом случае ширина каждой частотной полосы составит 40-50 Гц. Для получения достоверного усредненного значения спектральной плотности для каждой частотной полосы достаточно 5-10 значений. Отсюда, достаточное частотное разрешение при расчете энергетического спектра составляет (2) есть автокорреляционная функция случайного сигнала и представляет собой функцию спектральной плотности мощности случайного сигнала. Для стационарных эргодических процессов эта функция не случайна и остается постоянной для любой реализации случайного сигнала. Значения отдельных спектральных составляющих этой функции, вычисленные по любой из реализаций одного и того же случайного сигнала, имеют гораздо меньшие вариации, чем амплитуды тех же спектральных составляющих исходного сигнала. Наиболее приемлемым способом определения функции спектральной плотности мощности является числовой, поскольку аналоговыми методами получить автокорреляционную функцию случайного сигнала сложно. Однако при численном методе определения энергетического спектра случайного сигнала для очень коротких реализаций сигнала существенное значение имеют частота дискретизации исходного аналогового сигнала, величина шага квантования при расчете автокорреляционной функции и частотное разрешение при расчете по ней энергетического спектра [4]. При выборе этих величин необходимо руководствоваться необходимостью обеспечения достаточной достоверности получаемого спектра, т.е. минимальных различий спектров, получаемых по коротким реализациям одного и того же стационарного случайного процесса, и минимум затрат машинного времени, необходимого для анализа спектра. Выбор частоты дискретизации исходного сигнала обычно осуществляют в соответствии с теоремой Котельникова. Однако поскольку наиболее информативной частью спектра акустических шумов пчелиной семьи является диапазон от 100 до 600 Гц, в то время как частотный диапазон самих сигналов простирается до десятков кГц, то выбор частоты дискретизации здесь не является тривиальным вопросом. Если выбрать частоту дискретизации, ориентируясь на верхнюю границу интересующего нас частотного диапазона (fд = 1200-1500 Гц), то наличие в сигнале частот свыше 600 Гц приведет к дополнительному разбросу спектров отдельных реализаций одного и того же сигнала. С другой стороны, увеличение частоты дискретизации будет приводить к резкому увеличению объема вычислительных работ и необходимых ресурсов оперативной памяти ЭВМ, что нежелательно. Поэтому наилучшим решением является сохранение низкой частоты дискретизации (1200-1500 Гц) при предварительной частотной фильтрации исходного аналогового сигнала с выделением из него интересующего нас частотного диапазона (100-600 Гц). При этом дополнительного разброса спектров отдельных реализаций происходить не будет. Шаг квантования при расчете автокорреляционной функции обычно выбирается равным шагу дискретизации исходного сигнала. Шаг квантования по частоте при расчете энергетического спектра должен выбираться из следующих соображений. Для удобства анализа и надежного распознавания отдельных состояний пчелиной семьи по энергетическому спектру желательно разделить весь частотный диапазон на 10-12 полос и получать усредненные значения спектральной плотности для этих полос. В этом случае ширина каждой частотной полосы составит 40-50 Гц. Для получения достоверного усредненного значения спектральной плотности для каждой частотной полосы достаточно 5-10 значений. Отсюда, достаточное частотное разрешение при расчете энергетического спектра составляет 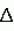 f = 5 Гц. Существенными отличительными признаками предлагаемого способа распознавания состояний пчелиной семьи по издаваемому ею акустическому шуму, использующему вычисление энергетического спектра короткой реализации сигнала, являются предварительная аналоговая частотная фильтрация анализируемого сигнала с выделением наиболее информативного частотного диапазона 100-600 Гц, что позволяет устранить дополнительный разброс спектральных составляющих, полученных по различным реализациям сигнала, при сохранении низкой частоты дискретизации; разделение всего анализируемого частотного диапазона энергетического спектра на небольшое число полос с вычислением усредненных для данных полос значений спектральной плотности, и идентификация различных состояний пчелиной семьи путем сопоставительного анализа усредненных значений спектральной плотности для этих полос. Кроме того, для повышения достоверности распознавания при очень коротких реализациях с целью исключения из анализа звуков помех, издаваемых отдельными пчелами, осуществляют выбор реализаций по критерию незначимых различий из дисперсий. Источники информации 1. Еськов Е. К. Акустическая сигнализация общественных насекомых. М.: Наука. - 1979. 2. Еськов Е.К. Управление процессами жизнедеятельности медоносных пчел и их оптимизация. М.: Всесоюзная академия сельскохозяйственных наук имени В.И. Ленина. - 1982. 3. Баскаков С. И. Радиотехнические цепи и сигналы. М.: Высшая школа - 1983. 4. Бендат Дж., Пирсол А., Измерение и анализ случайных процессов. Пер. с англ. М.: Мир - 1971. 5. Дьяконов В.П. Справочник по алгоритмам и программам на языке бейсик для персональных ЭВМ. - М.: Наука, главная редакция физико-математической литературы. - 1987. f = 5 Гц. Существенными отличительными признаками предлагаемого способа распознавания состояний пчелиной семьи по издаваемому ею акустическому шуму, использующему вычисление энергетического спектра короткой реализации сигнала, являются предварительная аналоговая частотная фильтрация анализируемого сигнала с выделением наиболее информативного частотного диапазона 100-600 Гц, что позволяет устранить дополнительный разброс спектральных составляющих, полученных по различным реализациям сигнала, при сохранении низкой частоты дискретизации; разделение всего анализируемого частотного диапазона энергетического спектра на небольшое число полос с вычислением усредненных для данных полос значений спектральной плотности, и идентификация различных состояний пчелиной семьи путем сопоставительного анализа усредненных значений спектральной плотности для этих полос. Кроме того, для повышения достоверности распознавания при очень коротких реализациях с целью исключения из анализа звуков помех, издаваемых отдельными пчелами, осуществляют выбор реализаций по критерию незначимых различий из дисперсий. Источники информации 1. Еськов Е. К. Акустическая сигнализация общественных насекомых. М.: Наука. - 1979. 2. Еськов Е.К. Управление процессами жизнедеятельности медоносных пчел и их оптимизация. М.: Всесоюзная академия сельскохозяйственных наук имени В.И. Ленина. - 1982. 3. Баскаков С. И. Радиотехнические цепи и сигналы. М.: Высшая школа - 1983. 4. Бендат Дж., Пирсол А., Измерение и анализ случайных процессов. Пер. с англ. М.: Мир - 1971. 5. Дьяконов В.П. Справочник по алгоритмам и программам на языке бейсик для персональных ЭВМ. - М.: Наука, главная редакция физико-математической литературы. - 1987.
ФОРМУЛА ИЗОБРЕТЕНИЯСпособ диагностирования состояний пчелиной семьи по издаваемому ею акустическому шуму, использующий численное разложение акустического сигнала путем вычисления энергетического спектра по автокорреляционной функции реализации дискретизованного сигнала, отличающийся тем, что исходный аналоговый сигнал предварительно подвергают частотной фильтрации с выделением наиболее информативного частотного диапазона, например 100 - 600 Гц, затем сигнал дискретизируют и из него выбирают 3 - 5 коротких реализаций длительностью 0,5 - 1,0 с, отвечающих условию незначимых различий их дисперсий, и по одной из них рассчитывают энергетический спектр, который делят на 10 - 12 частотных полос, для которых рассчитывают усредненные значения спектральной плотности и производят распознавание состояния пчелиной семьи путем сопоставительного анализа, усредненных значений спектральной плотности этих же полос частот для диагностируемого состояния пчелиной семьи.Популярные патенты: 2238970 Штамм mycelia sterilia лх-1-продуцент комплекса биологически активных веществ, обладающих рострегуляторными свойствами ... Штамм выделен из корневой системы облепихи. В качестве питательной среды используют, г/л: глюкозу, KH2PO4, K2HPO4, MgSO4, К2SO4, MnSO4, FeSO4, аспарагин.Основным его недостатком является сложность его получения в промышленном масштабе, штамм оказывает эффективность только в прикорневой зоне определенного растения-хозяина.Целью изобретения является расширение источников сырья из новых штаммов и повышение активности регулятора роста.Цель достигается тем, что из корней лоха выделен новый штамм Mycelia sterilia ЛХ-1. Штамм хранится в Всероссийской коллекции промышленных микроорганизмов, коллекционный номер ВКПМ F-862. Штамм предназначен для получения биостимулятора роста ... 2460269 Малогабаритный картофелеуборочный комбайн ... случаях угол наклона верхней ветви ворохоподъемного элеватора 15 регулируется перестановкой болта по отверстиям сектора 17.На верхней ветви клубнеприемного элеватора 11 клубни, встретившись с отражательным щитком 12, смещаются в сторону, сходят с поверхности элеватора и по лотку 24 поступают в тару (корзину, мешок). При этом мелкие почвенные примеси остаются на поверхности элеватора 11, проходят через зазор между нижней кромкой отражательного щитка 12 и прутками элеватора 11 и выбрасываются вперед на землю. В необходимых случаях этот зазор можно отрегулировать таким образом, чтобы через него проходили мелкие клубни.На сухих глинистых почвах клубни по лотку 24 сходят на землю, ... 2477044 Искусственная рыболовная приманка (варианты) ... корпуса гибким элементом при поклевке и для удобства рыболова при снятии рыбы с крючка. Гибкий элемент 6 в данном случае не является ограничителем растяжения корпуса приманки, а ограничителем 4 является деталь, закрепленная на элементе 6 в хвостовой части 2 корпуса. В качестве ограничителя в этом случае может использоваться крючок, жала которого утоплены в хвостовой части корпуса, или шумовой шарик, или капсула, или шарик с глазком или цветной капелькой, или капсула с ароматизатором, или любая другая деталь, не дающая корпусу растягиваться при поклевке. Приманка благодаря ограничителю 4 может использоваться многократно даже при появлении на ней укусов, порезов и т.д. Сквозной канал ... 2149547 Пневматический опрыскиватель ... клапаном 8. Под давлением воздуха рабочая жидкость поступает по питающей магистрали 10 к пневматическим распыливающим наконечникам 6. Количество поступающей жидкости регулируется краном-регулятором расхода рабочей жидкости 12. Распыливание химического препарата осуществляется внутри пневматического распыливающего наконечника 6. Рабочая жидкость, пройдя через патрубок 25, попадет в осевой канал 22, а затем по радиальным каналам 23 - к сопловым отверстиям 24. Воздух из воздухопроводов 16 поступает в кольцевое пространство, образованное корпусом 20 пневматического распыливающего наконечника и диспергирующим устройством 21. Часть воздуха попадает в сопловые отверстия 24, где ... 2455815 Самоходный универсальный комбайн для уборки картофеля и топинамбура ... SU 1384248 А01D 15/04 или SU 650541 А01D 15/04, также SU 1271411 A01L 19/02; или SU 1184499, А01D 15/00; или SU 893160 А01D 15/04.Известные самоходные картофелеуборочные комбайны, смотри учебное пособие Петрова Г.Д. и Карева Е.Б. Самоходные картофелеуборочные комбайны КСК-4, М., Агропромиздат, 1986, в частности: с.14 - комбайн КСК-4А-1, или с.26 - комбайн КСК-4, или с.35 - комбайн КСК-4А-1.В качестве прототипа предлагается принять самоходный картофелеуборочный комбайн КСК-4А-1, имеющий основные рабочие органы и сборочные единицы: приемную часть, включающую в себя лемех, выкапывающие диски, основной элеватор с сужающими шнеками, второй элеватор, выносной транспортер, ... |
Еще из этого раздела: 2171570 Устройство для группового учета надоев молока при доении 2248687 Способ весеннего боронования озимых культур и зубовая борона для его осуществления 2287923 Роторный энергосберегающий мостовой агрегат для сельскохозяйственных работ 2201244 Препарат для защиты животных и растений 2038763 Регулятор вакуума 2182889 Дезинфицирующее средство 2423036 Биоконтейнер для посадки растений 2482660 Способ выращивания рапса ярового на семена 2175177 Агромост с оснасткой для прокладки и уплотнения постоянных грунтовых колей 2432394 Ингибирование образования биогенного сульфида посредством комбинации биоцида и метаболического ингибитора |
Изобретения в сельском хозяйстве
Обработка почвы в сельском и лесном хозяйствах
Посадка, посев, удобрение
Уборка урожая, жатва
Обработка и хранение продуктов полеводства и садоводства
Садоводство, разведение овощей, цветов, риса, фруктов, винограда, лесное хозяйство
Новые виды растений или способы их выращивания
Производство молочных продуктов
Животноводство, разведение и содержание птицы, рыбы, насекомых, рыбоводство, рыболовство
Поимка, отлов или отпугивание животных
Консервирование туш животных, или растений или их частей
Биоцидная, репеллентная, аттрактантная или регулирующая рост растений активность химических соединений или препаратов
Хлебопекарные печи, машины и прочее оборудование для хлебопечения
Машины или оборудование для приготовления или обработки теста
Обработка муки или теста для выпечки, способы выпечки, мучные изделия
|
|
||