Понижающая регуляция экспрессии гена с помощью искусственных микрорнкПатент на изобретение №: 2492239 Автор: МАКГОНИДЖЛ Брайан (US) Патентообладатель: Е.И.ДЮПОН ДЕ НЕМУР ЭНД КОМПАНИ (US) Дата публикации: 27 Января, 2012 Начало действия патента: 17 Декабря, 2008 Адрес для переписки: 129090, Москва, ул.Б.Спасская, 25, стр.3, ООО "Юридическая фирма Городисский и Партнеры", пат.пов. А.В.Мицу, рег. 364 Изображения    Изобретение относится к области биохимии, в частности к выделенному фрагменту нуклеиновой кислоты для понижения экспрессии последовательности-мишени, по существу являющейся дезоксирибонуклеотидной последовательностью, представленной в SEQ ID NO: 11. Раскрыта рекомбинантная конструкция для понижения экспрессии последовательности-мишени, включающая указанный выделенный фрагмент нуклеиновой кислоты, а также клетка маиса для понижения экспрессии последовательности-мишени, включающая указанную рекомбинантную конструкцию. Описан способ отбора клеток, имеющих пониженный уровень экспрессии, включающий трансформацию клетки маиса указанной рекомбинантной конструкцией и отбор клетки (клеток), у которых уровень экспрессии последовательности-мишени понижен по сравнению с уровнем экспрессии гена-мишени в клетке маиса дикого типа. Изобретение позволяет понижать экспрессию последовательности-мишени в растении. 4 н.п. ф-лы, 13 табл., 12 пр. По данной заявке испрашивается приоритет предварительной заявки на патент США ОБЛАСТЬ ИЗОБРЕТЕНИЯ Область по данному изобретению касается, в целом, молекулярной биологии растений. В частности, она касается конструкций и способов для понижающей регуляции экспрессии последовательностей-мишеней. ПРЕДПОСЫЛКИ МикроРНК (миРНК) были впервые идентифицированы лишь несколько лет назад, но уже стало ясно, что они играют важную роль в регулировании генной активности. Эти некодирующие РНК из 20-22 нуклеотидов обладают способностью гибридизоваться посредством спаривания оснований со специфическими мРНК-мишенями и подавлять экспрессию этих транскриптов, опосредуя или РНК-расщепление или трансляционную репрессию. Последние исследования показали, что миРНК обладают важными функциями при развитии. В растениях, как показали, они контролируют ряд процессов развития, включающих период цветения, морфологию листа, полярность органов, морфологию цветка и развитие корня (рассмотрено Mallory и Vaucheret (2006) Nat Genet 38: S31-36). С учетом установленной регуляторной роли миРНК вполне вероятно, что они также вовлечены в контроль некоторых из основных признаков культур, таких как засухоустойчивость и устойчивость к болезням. МиРНК транскрибируются РНК полимеразой II как полиаденилированные и кэппированные сигналы, известные как прай-миРНК (первичная миРНК pri-miRNA). Такие прай-миРНК процессируются до более мелких транскриптов, известных как пре-миРНК (pre-miRNA), и такие предшественники обладают способностью формировать стабильныешпилечные структуры (рассмотрено Bartel (2004) Cell 116: 281-297; Jones-Rhoades MW, Bartel DP, Bartel B. MicroRNAS and their regulatory roles in plants. Annu Rev Plant Biol. 2006;57:19-53). Несмотря на то, что прай-миРНК процессируются до пре-миРНК с помощью Drosha в ядре, и Dicer расщепляет пре-миРНК в цитоплазме многоклеточных, созревание миРНК в растениях отличается от пути метаболизма у животных,поскольку растения лишены гомолога Drosha. Вместо этого фермент РНКаза III DICER-LIKE 1 (DCL1), который гомологичен животному Dicer, может обладать функцией Drosha в дополнение к его известной функции в шпилечном процессинге (Kurihara и Watanabe (2004) Proc Natl Acad Sci 101: 12753-12758). В Arabidopsis недавно были описаны искусственные миРНК (имиРНК, amiRNA), нацеленные на вирусные мРНК последовательности (Niu et al. (2006) Nature Biotechnology 24:1420-1428) или на эндогенные гены (Schwab et al. (2006) Plant Cell 18:1121-1133). Конструкция имиРНК может быть экспрессирована под контролем различных промоторов для изменения пространственного характера сайленсинга (Schwab et al. (2006) Plant Cell 18:1121-1133). Искусственные миРНК заменяют микроРНК и комплементарную ей последовательность, отмеченную штрихом, в предшественнике миРНК и замещают последовательности, которые нацелены на мРНК, подлежащую сайленсингу. Сайленсинг эндогенными миРНК можно обнаружить в ряде пространственных, временных и связанных с развитием паттернов экспрессии (Parizotto et al. (2007) Genes Dev 18:2237-2242; Alvarez et al. (2006) Plant Cell 18:1134-51). Искусственные миРНК можно сконструировать как для захвата, так и для расширения разнообразия и специфичности в паттернах сайленсинга. До сих пор не сообщалось о применении имиРНК в культурных растениях. WO 2004/009779, опубликованная 29 января 2004 г., раскрывает композиции и способы модулирования экспрессии генов в растениях. Заявка на патент США 2005/0138689 правопреемника заявителя, опубликованная 23 июня 2005 г., описывает миРНК и их применение в сайленсинге последовательности-мишени. КРАТКОЕ ОПИСАНИЕ СПИСКА ПОСЛЕДОВАТЕЛЬНОСТЕЙ Данное изобретение можно более полно понять из следующего подробного описания и сопровождающего Списка последовательностей, который составляет часть данной заявки. Описания последовательностей резюмируют Список последовательностей, приложенный к настоящему документу. Список последовательностей включает однобуквенные коды символов нуклеотидных последовательностей и одно- и трехбуквенные коды для аминокислот, как определено в стандартах IUPAC-IUB, раскрытых в Nucleic Acids Research 13:3021-3030 (1985) и в Biochemical Journal 219(2):345-373 (1984). SEQ ID NO:1-10 соответствуют праймерам, используемым для амплификации предшественников геномной микроРНК (миРНК) маиса. SEQ ID NO:11-15 соответствуют последовательностям предшественника миРНК маиса для 159c, 164h, 168a, 169r и 396h, соответственно. SEQ ID NO:16 соответствуют последовательности искусственной миРНК (имиРНК), используемой для сайленсинга транскрипта фитоен-десатуразы маиса (PDS). SEQ ID NO:17-21 соответствуют SEQ ID NO:22-26 соответствуют предшественникам имиРНК для 159c-PDS, 164h-PDS, 168a-PDS, 169r-PDS и 396h-PDS, соответственно. Эти предшественники, в случае если экспрессируются в маисе, управляют сайленсингом эндогенного транскрипта PDS. SEQ ID NO:27-30 соответствуют усеченным предшественникам имиРНК 169r-PDS-sht, 169r-PDS-med, 396h-PDS-sht и 396-PDS-med, соответственно. 169r-PDS предшественник (SEQ ID NO:25) был укорочен до 11% его длины (169r-PDS-sht, SEQ ID NO:27) и 35% его длины (169r-PDS-med, SEQ ID NO:28) по сравнению с 169r-PDS. 396h-PDS предшественник (SEQ ID NO:26) был укорочен до 18% его длины (396h-PDS-sht, SEQ ID NO:29) и 46% его длины (396h-PDS-med, SEQ ID NO:30) по сравнению с 396h-PDS. Все из усеченных предшественников содержали миРНК и последовательности, отмеченные штрихом. SEQ ID NO:31-34 соответствуют предшественникам имиРНК для 159c-FAD, 168a-FAD, 169r-FAD и 396h-FAD, соответственно. Эти предшественники, в случае если экспрессируются в маисе, управляют сайленсингом эндогенного fad2-1 (десатураза жирной кислоты, отвечающая за превращение олеиновой кислоты в линолевую кислоту) транскрипта. SEQ ID NO:35-38 соответствуют миРНК-мишени и последовательностям, отмеченным штрихом, для летальной пятнистости листьев. SEQ ID NO:39-41 соответствуют миРНК-мишени и последовательностям, отмеченным штрихом, для белка устойчивости к нескольким лекарственным средствам, который является транспортным белком. SEQ ID NO:42-43 соответствуют последовательностям предшественников имиРНК для 168a-MRP и 396h-MRP, соответственно. Эти предшественники, в случае если экспрессируются в маисе, управляют сайленсингом MRP, что приводит к пониженным уровням фитиновой кислоты. КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ Данное изобретение относится к выделенному фрагменту нуклеиновой кислоты, включающему предшественник миРНК, причем указанный предшественник миРНК по существу соответствует дезоксирибонуклеотидной последовательности, представленной в SEQ ID NO:11, (i) где нуклеотиды 430-450 в SEQ ID NO:11 замещены первой вариабельной нуклеотидной субпоследовательностью, размер которой варьирует от примерно 19 до примерно 30 нуклеотидов в зависимости от последовательности-мишени, экспрессия которой подлежит понижению, и (ii) кроме того, где нуклеотиды 244-264 в SEQ ID NO:11 замещены второй вариабельной нуклеотидной субпоследовательностью, размер которой варьирует от примерно 19 до примерно 30 нуклеотидов, причем указанная вторая вариабельная нуклеотидная субпоследовательность способна гибридизоваться с первой вариабельной субпоследовательностью предшественника миРНК. Другие выделенные фрагменты нуклеиновых кислот, которые также представляют интерес, включают следующее: a) выделенный фрагмент нуклеиновой кислоты, включающий предшественник миРНК, причем указанный предшественник миРНК по существу соответствует дезоксирибонуклеотидной последовательности, представленной в SEQ ID NO:12, (i) где нуклеотиды 94-114 в SEQ ID NO:12 замещены первой вариабельной нуклеотидной субпоследовательностью, размер которой варьирует от примерно 19 до примерно 30 нуклеотидов в зависимости от последовательности-мишени, экспрессия которой подлежит понижению, и (ii) кроме того, где нуклеотиды 163-183 в SEQ ID NO:12 замещены второй вариабельной нуклеотидной субпоследовательностью, размер которой варьирует от примерно 19 до примерно 30 нуклеотидов, причем указанная вторая вариабельная нуклеотидная субпоследовательность способна гибридизоваться с первой вариабельной субпоследовательностью предшественника миРНК; b) выделенный фрагмент нуклеиновой кислоты, включающий предшественник миРНК, причем указанный предшественник миРНК по существу соответствует дезоксирибонуклеотидной последовательности, представленной в SEQ ID NO:13, (i) где нуклеотиды 53-73 в SEQ ID NO:13 замещены первой вариабельной нуклеотидной субпоследовательностью, размер которой варьирует от примерно 19 до примерно 30 нуклеотидов в зависимости от последовательности-мишени, экспрессия которой подлежит понижению, и (ii) кроме того, где нуклеотиды 97-117 в SEQ ID NO:13 замещены второй вариабельной нуклеотидной субпоследовательностью, размер которой варьирует от примерно 19 до примерно 30 нуклеотидов, причем указанная вторая вариабельная нуклеотидная субпоследовательность способна гибридизоваться с первой вариабельной субпоследовательностью предшественника миРНК; c) выделенный фрагмент нуклеиновой кислоты, включающий предшественник миРНК, причем указанный предшественник миРНК по существу соответствует дезоксирибонуклеотидной последовательности, представленной в SEQ ID NO:14, (i) где нуклеотиды 110-130 в SEQ ID NO:14 замещены первой вариабельной нуклеотидной субпоследовательностью, размер которой варьирует от примерно 19 до примерно 30 нуклеотидов в зависимости от последовательности-мишени, экспрессия которой подлежит понижению, и (ii) кроме того, где нуклеотиды 184-203 в SEQ ID NO:14 замещены второй вариабельной нуклеотидной субпоследовательностью, размер которой варьирует от примерно 19 до примерно 30 нуклеотидов, причем указанная вторая вариабельная нуклеотидная субпоследовательность способна гибридизоваться с первой вариабельной субпоследовательностью предшественника миРНК; и d) выделенный фрагмент нуклеиновой кислоты, включающий предшественник миРНК, причем указанный предшественник миРНК по существу соответствует дезоксирибонуклеотидной последовательности, изложенной в SEQ ID NO:15, (i) где нуклеотиды 83-103 в SEQ ID NO:15 замещены первой вариабельной нуклеотидной субпоследовательностью, размер которой варьирует от примерно 19 до примерно 30 нуклеотидов в зависимости от последовательности-мишени, экспрессия которой подлежит понижению, и (ii) кроме того, где нуклеотиды 172-192 в SEQ ID NO:15 замещены второй вариабельной нуклеотидной субпоследовательностью, размер которой варьирует от примерно 19 до примерно 30 нуклеотидов, причем указанная вторая вариабельная нуклеотидная субпоследовательность способна гибридизоваться с первой вариабельной субпоследовательностью предшественника миРНК. Любой из этих выделенных фрагментов нуклеиновой кислоты может быть использован для создания рекомбинантной конструкции, включающей эти выделенные фрагменты нуклеиновой кислоты, функционально связанные, по меньшей мере, с одной регуляторной последовательностью. Эти конструкции могут быть трансформированы в растительную клетку так, что трансформированная растительная клетка будет включать рекомбинантную конструкцию в своем геноме. В другом аспекте данное изобретение относится к способу понижения экспрессии гена-мишени в растительной клетке, включающему: (a) трансформацию, по меньшей мере, одной растительной клетки конструкцией нуклеиновой кислоты, включающей любой из выделенных фрагментов нуклеиновой кислоты, описанных в данном документе; и (b) отбор той трансформированной растительной клетки (клеток), у которой уровень экспрессии последовательности-мишени понижен по сравнению с уровнем экспрессии гена-мишени в растительной клетке дикого типа. ДЕТАЛЬНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ Информацию, соответствующую данной заявке, можно найти в заявках на патенты США Другие ссылки, которые можно использовать для понимания данного изобретения, включают заявку на патент США Раскрытие каждой ссылки, изложенной в данном документе, настоящим включено ссылкой во всей своей полноте. В данном документе и в приложенной Формуле изобретения формы единственного числа включают ссылки на множественное число, если в контексте четко не диктуется иное. Таким образом, например, ссылка на "растение" включает множество таких растений, ссылка на "клетку" включает одну или более клеток и их эквивалентов, известных специалисту в данной области, и т.д. В контексте данного раскрытия используется ряд выражений и аббревиатур. Представлены следующие определения.
Выражение "геном" означает следующее: (1) полный комплемент генетического материала (гены и некодирующие последовательности), присутствующего в каждой клетке организма, или в вирусе, или в органелле; (2) целый набор хромосом, наследованный как (гаплоидная) единица от одного родителя. "Потомство" включает любое последующее поколение растения. Потомство будет наследовать и стабильно сегрегировать гены и трансгены от своего родительского растения (растений). Единицы, приставки и символы могут быть выражены в приемлемой форме СИ (международная система единиц). Если не указано иное, нуклеиновые кислоты пишутся слева направо в 5'-3' ориентации; аминокислотные последовательности пишутся слева направо в ориентации амино-карбоксил, соответственно. Числовые диапазоны, перечисляемые в спецификации, охватывают числа, определяющие диапазон, и включают каждое целое число в определенном диапазоне. Аминокислоты могут быть обозначены в данном документе либо общеизвестными трехбуквенными символами, либо однобуквенными символами, рекомендованными Комиссией по биохимической номенклатуре IUPAC-IUB. Нуклеотиды аналогичным образом могут быть обозначены их общепринятыми однобуквенными кодами. Если иное не предусмотрено, выражения программного обеспечения, электрические и электроники, используемые в данном документе, определены в The New IEEE Standard Dictionary of Electrical and Electronics Terms (5th edition, 1993). Выражения, определенные ниже, более полно определены ссылкой на спецификацию в целом. Выражения Такая конструкция может включать любую комбинацию дезоксирибонуклеотидов, рибонуклеотидов и/или модифицированных нуклеотидов. Конструкция может транскрибироваться с образованием РНК, где РНК может быть способной формировать двухцепочечную РНК и/или шпилечную структуру. Такая конструкция может быть экспрессирована в клетке, или выделена, или получена синтетически. Конструкция может дополнительно включать промотор или другие последовательности, которые облегчают манипуляцию или экспрессию конструкции. В данном документе В данном документе В данном документе В данном документе "гетерологичная" по отношению к последовательности означает последовательность, которая происходит от чужеродных видов, или, если от одних и тех же видов, является по существу модифицированной преднамеренным вмешательством человека от ее нативной формы в композиции и/или геномном локусе. Например, по отношению к нуклеиновой кислоте это может быть нуклеиновая кислота, которая происходит от чужеродных видов, или сконструирована синтетически, или если от одних и тех же видов является по существу модифицированным преднамеренным вмешательством человека от ее нативной формы в композиции и/или геномном локусе. Гетерологичный белок может происходить от чужеродных видов или, если от одних и тех же видов, является по существу модифицированным преднамеренным вмешательством человека от своей первоначальной формы. Выражение "клетка-хозяин" означает клетку, которая включает или в которую введена конструкция нуклеиновой кислоты и которая поддерживает репликацию и/или экспрессию конструкции. Клетки-хозяева могут быть прокариотическими клетками, такими как E. coli, или эукариотическими клетками, такими как клетки грибов, дрожжей, насекомого, амфибии, нематоды или млекопитающего. Альтернативно, клетки-хозяева являются растительными клетками однодольных или двудольных растений. Примером клетки-хозяина однодольного растения является клетка-хозяин маиса. Выражение Выражение "орган растения" означает растительную ткань или группу тканей, которая составляет морфологически и функционально отдельную часть растения. Выражение Выражение Выражение "выделенный" означает материал, такой как нуклеиновая кислота или белок, который является: (1) по существу или существенно не содержащим компоненты, которые обычно сопровождают или взаимодействуют с материалом, как его обнаруживают в его естественной среде, или (2) если материал в своей естественной среде, материалом, который изменен преднамеренным вмешательством человека в композицию и/или помещен в локус в клетке, отличного от локуса, нативного данному материалу. В данном документе РНК интерференция означает процесс специфического по последовательности пост-транскрипционного генного сайленсинга у животных, опосредованного короткими интерферирующими РНК (киРНК) (Fire et al., Nature 391:806 1998). Соответствующий процесс у растений обычно называют пост-транскрипционным генным сайленсингом (PTGS) или РНК сайленсингом, а также называют подавлением в грибах. Процесс пост-транскрипционного генного сайленсинга считается эволюционно-консервативным клеточным защитным механизмом, используемым для предотвращения экспрессии чужеродных генов, и обычно совместно используется разнообразной флорой и таксономическими типами (Fire et al., Trends Genet. 15:358 1999). Такая защита от экспрессии чужеродных генов могла развиться в ответ на продуцирование двухцепочечных РНК (днРНК), полученных от вирусной инфекции или от случайной интеграции транспозонных элементов в геном хозяина, путем клеточного ответа, который специфически разрушает гомологичную одноцепочечную РНК вирусной геномной РНК. Присутствие двухцепочечной РНК в клетках вызывает ответ РНК-интерференции через механизм, который полностью еще не охарактеризован. Присутствие длинных днРНК в клетках стимулирует активность фермента рибонуклеаза III, названного РНК-интерференция исследована на ряде систем. Fire et al. (Nature 391:806 1998) первыми наблюдали РНК-интерференцию у C. elegans. Wianny и Goetz (Nature Cell Biol. 2:70 1999) описывают РНК-интерференцию, опосредованную двухцепочечной РНК в зародышах мышей. Hammond et al. (Nature 404:293 2000) описывают РНК-интерференцию в клетках Drosophila, трансфицированных двухцепочечной РНК. Elbashir et al. (Nature 411:494 2001) описывают РНК-интерференцию, индуцированную введением дуплексов синтетических 21-нуклеотидных РНК в культивируемых клетках млекопитающего, включая первичную почку человека и HeLa клетки. Малые РНК играют важную роль в регуляции экспрессии гена. Регуляция многих связанных с развитием процессов, включая цветение, контролируется малыми РНК. Теперь можно конструировать изменения в генной экспрессии растительных генов путем использования трансгенных конструкций, которые продуцируют малые РНК в растении. Малые РНК, как оказывается, функционируют спариванием оснований с комплементарной РНК или ДНК последовательностями-мишенями. При связывании с РНК малые РНК запускают или расщепление РНК, или трансляционное ингибирование последовательности-мишени. При связывании с ДНК последовательностями-мишенями считается, что малые РНК могут опосредовать ДНК метилирование последовательности-мишени. Следствием этих событий независимо от специфического механизма является то, что ингибируется генная экспрессия. МикроРНК (миРНК) являются некодирующими РНК длиной от примерно 19 до примерно 24 нуклеотидов (нт), которые идентифицированы и у животных, и у растений (Lagos-Quintana et al., Science 294:853-858 2001, Lagos-Quintana et al., Curr. Biol. 12:735-739 2002; Lau et al., Science 294:858-862 2001; Lee и Ambros, Science 294:862-864 2001; Llave et al., Plant Cell 14:1605-1619 2002; Mourelatos et al., Genes. Dev. 16:720-728 2002; Park et al., Curr. Biol. 12:1484-1495 2002; Reinhart et al., Genes. Dev. 16:1616-1626 2002). Они получены из более длинных транскриптов-предшественников, размер которых варьирует от приблизительно 70 до 200 нт, и эти транскрипты-предшественники обладают способностью формировать стабильные шпилечные структуры. У животных фермент, вовлеченный в процессинг предшественников миРНК, называют Dicer, РНКаза III-подобный белок (Grishok et al., Cell 106:23-34 2001; Hutvagner et al., Science 293:834-838 2001; Ketting et al., Genes. Dev. 15:2654-2659 2001). У растений также есть Dicer-подобный фермент, DCL1 (ранее называемый CARPEL FACTORY/SHORT INTEGUMENTS1/ SUSPENSOR1), и последние данные показывают, что он, подобно Dicer, вовлечен в процессинг шпилечных предшественников для образования зрелых миРНК (Park et al., Curr. Biol. 12:1484-1495 2002; Reinhart et al., Genes. Dev. 16:1616-1626 2002). Кроме того, из последней работы становится ясно, что, по меньшей мере, некоторые шпилечные предшественники миРНК образуются как более длинные полиаденилированные транскрипты, и некоторые другие миРНК и связанные шпильки могут присутствовать в отдельном транскрипте (Lagos-Quintana et al., Science 294:853-858 2001; Lee et al., EMBO J 21:4663-4670 2002). Последняя работа также рассматривала выбор цепи миРНК от продукта двухцепочечной РНК, происходящего от процессинга шпильки с помощью DICER (Schwartz et al., 2003, Cell 115:199-208). Становится ясно, что стабильность (т.е. содержание G:C против A:U и/или ошибочные спаривания) двух концов процессированной двухцепочечной РНК влияет на выбор цепи, при этом конец с низкой стабильностью легче раскрутить геликазной активностью. Цепь с 5' концом низкой стабильности включена в RISC комплекс, тогда как другая цепь распадается. У животных существуют прямые доказательства роли специфических миРНК в развитии. Обнаружили, что lin-4 и let-7 миРНК у C. elegans контролируют временное развитие на основе фенотипов, полученных при мутировании генов, продуцирующих lin-4 и let-7 миРНК (Lee et al., Cell 75:843-854 1993; Reinhart et al., Nature 403-901-906 2000). Кроме того, обе миРНК отображают временной паттерн экспрессии в соответствии с их ролями во временном паттерне развития. Другие животные миРНК отображают регулируемые развитием паттерны экспрессии как временные, так и специфические для ткани (Lagos-Quintana et al., Science 294:853-853 2001, Lagos-Quintana et al., Curr. Biol. 12:735-739 2002; Lau et al., Science 294:858-862 2001; Lee и Ambros, Science 294:862-864 2001), что привело к гипотезе, что миРНК могут быть во многих случаях вовлечены в регуляцию важных процессов развития. Кроме того, в растениях дифференциальные паттерны экспрессии многих миРНК предполагают роль в развитии (Llave et al., Plant Cell 14:1605-1619 2002; Park et al., Curr. Biol. 12:1484-1495 2002; Reinhart et al., Genes. Dev. 16:1616-1626 2002). Тем не менее, роль в развитии для миРНК непосредственно не доказана в растениях, поскольку до настоящего времени не было никаких сообщений об относящемся к развитию фенотипе, связанном со специфической растительной миРНК. По-видимому, миРНК регулируют гены-мишени связыванием с комплементарными последовательностями, расположенными в транскриптах, которые продуцируются этими генами. В случае lin-4 и let-7 сайты-мишени расположены в 3' UTR мРНК-мишеней (Lee et al., Cell 75:843-854 1993; Wightman et al., Cell 75:855-862 1993; Reinhart et al., Nature 403:901-906 2000; Slack et al., Mol. Cell 5:659-669 2000), и имеется несколько ошибочных спариваний между lin-4 и let-7 миРНК и их сайтами-мишенями. По-видимому, связывание lin-4 или let-7 миРНК вызывает понижающую регуляцию установившихся уровней белка, кодируемого мРНК-мишенью, не влияя на сам транскрипт (Olsen и Ambros, Dev. Biol. 216:671-680 1999). С другой стороны, последние данные указывают на то, что миРНК могут в некоторых случаях вызывать специфическое РНК расщепление транскрипта-мишени на сайте-мишени (Hutvagner и Zamore, Science 297:2056-2060 2002; Llave et al., Plant Cell 14:1605-1619 2002). Вероятно, что миРНК могут вступать, по меньшей мере, в два пути метаболизма регуляции гена-мишени: понижающую регуляцию белка и РНК расщепление. МикроРНК, вступающие в путь метаболизма РНК расщепления, включены в комплекс РНК-индуцированного сайленсинга (RISC), который подобен или идентичен тому, что наблюдался для РНК-интерференции. Данное изобретение касается выделенного фрагмента нуклеиновой кислоты, включающего предшественник миРНК, причем указанный предшественник миРНК по существу соответствует дезоксирибонуклеотидной последовательности, изложенной в SEQ ID NO:11, (i) где нуклеотиды 430-450 в SEQ ID NO:11 замещены первой вариабельной нуклеотидной субпоследовательностью, размер которой варьирует от примерно 19 до примерно 30 нуклеотидов в зависимости от последовательности-мишени, экспрессия которой подлежит понижению, и (ii) кроме того, где нуклеотиды 244-264 в SEQ ID NO:11 замещены второй вариабельной нуклеотидной субпоследовательностью, размер которой варьирует от примерно 19 до примерно 30 нуклеотидов, причем указанная вторая вариабельная нуклеотидная субпоследовательность способна гибридизоваться с первой вариабельной субпоследовательностью предшественника миРНК. Этот выделенный фрагмент нуклеиновой кислоты, включающий предшественник миРНК, также можно называть Другие выделенные нуклеиновые фрагменты интереса включают следующее: a) транскрибированный из выделенного фрагмента нуклеиновой кислоты, включающий предшественник миРНК, причем указанный предшественник миРНК по существу соответствует дезоксирибонуклеотидной последовательности, изложенной в SEQ ID NO:12, (i) где нуклеотиды 94-114 в SEQ ID NO:12 замещены первой вариабельной нуклеотидной субпоследовательностью, размер которой варьирует от примерно 19 до примерно 30 нуклеотидов в зависимости от последовательности-мишени, экспрессия которой подлежит понижению, и (ii) кроме того, где нуклеотиды 163-183 в SEQ ID NO:12 замещены второй вариабельной нуклеотидной субпоследовательностью, размер которой варьирует от примерно 19 до примерно 30 нуклеотидов, причем указанная вторая вариабельная нуклеотидная субпоследовательность способна гибридизоваться с первой вариабельной субпоследовательностью предшественника миРНК; b) выделенный фрагмент нуклеиновой кислоты, включающий предшественник миРНК, причем указанный предшественник миРНК по существу соответствует дезоксирибонуклеотидной последовательности, изложенной в SEQ ID NO:13, (i) где нуклеотиды 53-73 в SEQ ID NO:13 замещены первой вариабельной нуклеотидной субпоследовательностью, размер которой варьирует от примерно 19 до примерно 30 нуклеотидов в зависимости от последовательности-мишени, экспрессия которой подлежит понижению, и (ii) кроме того, где нуклеотиды 97-117 в SEQ ID NO:13 замещены второй вариабельной нуклеотидной субпоследовательностью, размер которой варьирует от примерно 19 до примерно 30 нуклеотидов, причем указанная вторая вариабельная нуклеотидная субпоследовательность способна гибридизоваться с первой вариабельной субпоследовательностью предшественника миРНК; c) выделенный фрагмент нуклеиновой кислоты, включающий предшественник миРНК, причем указанный предшественник миРНК по существу соответствует дезоксирибонуклеотидной последовательности, изложенной в SEQ ID NO:14, (i) где нуклеотиды 110-130 в SEQ ID NO:14 замещены первой вариабельной нуклеотидной субпоследовательностью, размер которой варьирует от примерно 19 до примерно 30 нуклеотидов в зависимости от последовательности-мишени, экспрессия которой подлежит понижению, и (ii) кроме того, где нуклеотиды 184-203 в SEQ ID NO:14 замещены второй вариабельной нуклеотидной субпоследовательностью, размер которой варьирует от примерно 19 до примерно 30 нуклеотидов, причем указанная вторая вариабельная нуклеотидная субпоследовательность способна гибридизоваться с первой вариабельной субпоследовательностью предшественника миРНК; и d) выделенный фрагмент нуклеиновой кислоты, включающий предшественник миРНК, причем указанный предшественник миРНК по существу соответствует дезоксирибонуклеотидной последовательности, изложенной в SEQ ID NO:15, (i) где нуклеотиды 83-103 в SEQ ID NO:15 замещены первой вариабельной нуклеотидной субпоследовательностью, размер которой варьирует от примерно 19 до примерно 30 нуклеотидов в зависимости от последовательности-мишени, экспрессия которой подлежит понижению, и (ii) кроме того, где нуклеотиды 172-192 в SEQ ID NO:15 замещены второй вариабельной нуклеотидной субпоследовательностью, размер которой варьирует от примерно 19 до примерно 30 нуклеотидов, причем указанная вторая вариабельная нуклеотидная субпоследовательность способна гибридизоваться с первой вариабельной субпоследовательностью предшественника миРНК. Любой из этих выделенных фрагментов нуклеиновой кислоты можно использовать для создания рекомбинантной конструкции, включающей эти выделенные фрагменты нуклеиновой кислоты, функционально связанные, по меньшей мере, с одной регуляторной последовательностью. Такие конструкции можно трансформировать в растительную клетку так, что трансформированная растительная клетка будет включать рекомбинантную конструкцию в своем геноме. Предпочтительно, растительная клетка может быть растительной клеткой однодольного растения. Примеры растительных клеток однодольных растений включают, но без ограничения, клетки маиса, сорго, пшеницы, риса, овса, ржи, ячменя, сахарного тростника, просо, бамбука, банана и орхидных. Наиболее предпочтительной растительной клеткой однодольного растения является клетка маиса. В другом аспекте данное изобретение касается способа понижения экспрессии последовательности-мишени в растительной клетке, причем указанный способ включает: (a) осуществление трансформации, по меньшей мере, одной растительной клетки конструкцией нуклеиновой кислоты, включающей любой из выделенных фрагментов нуклеиновой кислоты, описанный в данном документе; и (b) отбор такой трансформированной растительной клетки (клеток), у которых уровень экспрессии последовательности-мишени понижен по сравнению с уровнем экспрессии последовательности-мишени в растительной клетке дикого типа. Биоинформационные подходы успешно используются для прогнозирования мишеней для растительных миРНК (Llave et al., Plant Cell 14:1605-1619 2002; Park et al., Curr. Biol. 12:1484-1495 2002; Rhoades et al., Cell 110:513-520 2002) и, таким образом, по-видимому, растительные миРНК имеют более высокую общую комплементарность с их предполагаемыми мишенями, чем животные миРНК. Большинство из таких предсказанных транскриптов-мишеней растительных миРНК кодируют членов семейств транскрипционных факторов, вовлеченных в связанное с развитием структурирование растения или клеточную дифференциацию. Общие категории последовательностей интереса включают, например, гены, вовлеченные в регуляцию или информацию, такие как цинковые пальцы, транскрипционные факторы, гомеозисные гены или модуляторы клеточного цикла и клеточной смерти, вовлеченные в коммуникацию, такие как киназы, и вовлеченные в осуществление общих для клеток функций, такие как белки теплового шока. Последовательности-мишени могут включать кодирующие участки и некодирующие участки, такие как промоторы, энхансеры, терминаторы, интроны и подобное. Последовательность-мишень может быть эндогенной последовательностью или может быть введенной гетерологичной последовательностью, или трансгеном. Например, способы можно использовать для изменения регуляции или экспрессии трансгена, или для удаления трансгена или другой введенной последовательности, такой как введенный сайт сайт-специфической рекомбинации. Последовательность-мишень также может быть последовательностью из патогена, например последовательность-мишень может быть из растительного патогена, такого как вирус, плесень или грибок, насекомое или нематода. МиРНК может быть экспрессирована в растении, которое при инфекции или заражении будет нацелена на патоген и будет придавать некоторую степень устойчивости растению. В растениях другие категории последовательностей-мишеней включают гены, влияющие на агрономические признаки, устойчивость к насекомым, устойчивость к болезням, устойчивость к гербицидам, стерильность, характеристики зерна и коммерческие продукты. Гены интереса также включают вовлеченные в метаболизм масла, крахмала, углевода или питательных веществ, а также влияющие, например, на размер зерна, наполнение сахарозой и подобное. Качество зерна отражается в признаках, таких как уровни и типы масел, насыщенных и ненасыщенных, качество и количество незаменимых аминокислот и уровни целлюлозы. Например, гены биосинтетического пути метаболизма фитиновой кислоты могут быть супрессированы для образования фенотипа высокой доступности фосфора. См., например, ферменты биосинтеза фитиновой кислоты, включающие полинуклеотиды инозитол-полифосфат-киназы-2, раскрытые в WO 02/059324, полинуклеотиды инозитол-1,3,4-трифосфат-5/6-киназы, раскрытые в WO 03/027243, и полинуклеотиды мио-инозитол-1-фосфат-синтазы и другие полинуклеотиды биосинтеза фитатов, раскрытые в WO 99/05298, все из которых включены ссылкой в данный документ. Гены в пути легнификации могут быть супрессированы для усиления перевариваемости или доступности энергии. Гены, влияющие на клеточный цикл или клеточную смерть, могут быть супрессированы для влияния на рост или стрессовый ответ. Гены, влияющие на ДНК репарацию и/или рекомбинацию, могут быть супрессированы для повышения генетической вариабельности. Могут быть супрессированы гены, влияющие на период цветения, а также гены, влияющие на фертильность. Любая последовательность-мишень может быть супрессирована для оценки или подтверждения ее роли в конкретном признаке или фенотипе, или для анализа молекулярного, регуляторного, биохимического или протеомного пути метаболизма или сети. Можно использовать ряд промоторов. Эти промоторы можно выбрать в зависимости от желаемого результата. Следует признать, что различные применения будут расширены применением различных промоторов в растительных экспрессирующих кассетах для модуляции временного паттерна, расположения и/или уровня экспрессии миРНК. Такие растительные экспрессирующие кассеты также могут включать, при желании, промоторный регуляторный участок (например, одна обсуждаемая индуцируемая, конститутивная, экологически или в зависимости от развития регулируемая или клетка- или ткане-специфическая/селективная экспрессия), сайт начала инициации транскрипции, сайт связывания с рибосомой, сигнал процессинга РНК, сайт терминации транскрипции и/или сигнал полиаденилирования. Можно применять конститутивные, предпочтительные для ткани или индуцируемые промоторы. Примеры конститутивных промоторов включают 35S участок инициации транскрипции вируса мозаики цветной капусты (CaMV), 1'- или 2'-промотор, полученный от T-ДНК Agrobacterium tumefaciens, ubiquitin 1 промотор, Smas промотор, промотор дегидрогеназы коричного спирта (патент США Примерами индуцируемых промоторов являются Adh1 промотор, который индуцируется гипоксией или холодовым стрессом, Hsp70 промотор, который индуцируется тепловым стрессом, PPDK промотор и пепкарбоксилазы промотор, оба индуцируемые светом. Также используются промоторы, которые индуцируются химически, такие как In2-2 промотор, который индуцируется сафенером (патент США Примеры промоторов под связанным с развитием контролем включают промоторы, которые инициируют транскрипцию, предпочтительно в определенных тканях, таких как листья, корни, плод, семена или цветки. Иллюстративным промотором является специфический для пыльника промотор 5126 (патенты США В некоторых вариантах осуществления будет полезным экспрессировать ген от индуцируемого промотора, особенно от патоген-индуцируемого промотора. Такие промоторы включают промоторы от связанных с патогенезом белков (PR белки), которые индуцируются после инфекции патогеном; например, PR белки, SAR белки, бета-1,3-глюканаза, хитиназа и т.д. См., например, Redolfi et al. (1983) Neth. J. Plant Pathol. 89:245-254; Uknes et al. (1992) Plant Cell 4:645-656; и Van Loon (1985) Plant Mol. Virol. 4:111-116. См. также WO 99/43819, включенную в данный документ ссылкой. Представляют интерес промоторы, экспрессируемые локально в или рядом с сайтом патогенной инфекции. См., например, Marineau et al. (1987) Plant Mol. Biol. 9:335-342; Matton et al. (1989) Molecular Plant-Microbe Interactions 2:325-331; Somsisch et al. (1986) Proc. Natl. Acad. Sci. USA 83:2427-2430; Somsisch et al. (1988) Mol. Gen. Genet. 2:93-98; и Yang (1996) Proc. Natl. Acad. Sci. USA 93:14972-14977. См. также, Chen et al. (1996) Plant J. 10:955-966; Zhang et al. (1994) Proc. Natl. Acad. Sci. USA 91:2507-2511; Warner et al. (1993) Plant J. 3:191-201; Siebertz et al. (1989) Plant Cell 1:961-968; патент США Кроме того, так как патогены находят вход в растения через раны или повреждение насекомым, в конструкциях полинуклеотидов можно использовать индуцируемый раной промотор. Такие индуцируемые раной промоторы включают ген ингибитора протеиназы (pin II) картофеля (Ryan (1990) Ann. Rev. Phytopath. 28:425-449; Duan et al. (1996) Nature Biotech. 14:494-498); wun1 и wun2, патент США Химически регулируемые промоторы можно использовать для модуляции экспрессии гена в растении путем применения экзогенного химического регулятора. В зависимости от цели промотор может быть химически индуцируемым промотором, где применение химиката индуцирует экспрессию гена, или химически репрессируемым промотором, где применение химиката репрессирует экспрессию гена. Химически индуцируемые промоторы известны в данной области и включают, но без ограничения, In2-2 промотор маиса, который активируется сафенерами бензолсульфонамидных гербицидов, GST промотор маиса, который активируется гидрофобными электрофильными соединениям, используемыми как довсходовые гербициды, и PR-1a промотор табака, который активируется салициловой кислотой. Другие химически регулируемые промоторы интереса включают реагирующие на стероиды промоторы (см., например, индуцируемый глюкокортикоидом промотор у Schena et al. (1991) Proc. Natl. Acad. Sci. USA 88:10421-10425 и McNellis et al. (1998) Plant J. 14(2):247-257), индуцируемый тетрациклином и репрессируемый тетрациклином промоторы (см., например, Gatz et al. (1991) Mol. Gen. Genet. 227:229-237 и патенты США Предпочтительные для ткани промоторы можно применять для мишеневой усиленной экспрессии последовательности интереса в конкретной растительной ткани. Предпочтительные для ткани промоторы включают Yamamoto et al. (1997) Plant J. 12(2):255-265; Kawamata et al. (1997) Plant Cell Physiol. 38(7):792-803; Hansen et al. (1997) Mol. Gen Genet. 254(3):337-343; Russell et al. (1997) Transgenic Res. 6(2):157-168; Rinehart et al. (1996) Plant Physiol. 112(3):1331-1341; Van Camp et al. (1996) Plant Physiol. 112(2):525-535; Canevascini et al. (1996) Plant Physiol. 112(2):513-524; Yamamoto et al. (1994) Plant Cell Physiol. 35(5):773-778; Lam (1994) Results Probl. Cell Differ. 20:181-196; Orozco et al. (1993) Plant Mol Biol. 23(6):1129-1138; Matsuoka et al. (1993) Proc Natl. Acad. Sci. USA 90(20):9586-9590; и Guevara-Garcia et al. (1993) Plant J. 4(3):495-505. Такие промоторы могут быть модифицированы, если необходимо, для слабой экспрессии. Предпочтительные для листьев промоторы известны в данной области. См., например, Yamamoto et al. (1997) Plant J. 12(2):255-265; Kwon et al. (1994) Plant Physiol. 105:357-67; Yamamoto et al. (1994) Plant Cell Physiol. 35(5):773-778; Gotor et al. (1993) Plant J. 3:509-18; Orozco et al. (1993) Plant Mol. Biol. 23(6):1129-1138; и Matsuoka et al. (1993) Proc. Natl. Acad. Sci. USA 90(20):9586-9590. Кроме того, также можно использовать промоторы cab и rubisco. См., например, Simpson et al. (1958) EMBO J 4:2723-2729 и Timko et al. (1988) Nature 318:57-58. Предпочтительные для корней промоторы известны и могут быть выбраны из многих, доступных из литературы или выделенных de novo из различных совместимых видов. См., например, Hire et al. (1992) Plant Mol. Biol. 20(2):207-218 (специфический для корня сои ген глютаминсинтетазы); Keller и Baumgartner (1991) Plant Cell 3(10):1051-1061 (специфический для корня элемент управления в GRP 1.8 гене фасоли обыкновенной); Sanger et al. (1990) Plant Mol. Biol. 14(3):433-443 (специфический для корня промотор гена маннопинсинтазы (MAS) Agrobacterium tumefaciens); и Miao et al. (1991) Plant Cell 3(1):11-22 (кДНК клон полной длины, кодирующий цитозольную глутаминсинтетазу (GS), который экспрессируется в корнях и корневых клубеньках сои). См. также Bogusz et al. (1990) Plant Cell 2(7):633-641, где описаны два специфических для корня промотора, выделенных из генов гемоглобина фиксирующего азот небобового Parasponia andersonii и родственного нефиксирующего азот небобового Trema tomentosa. Промоторы этих генов были связаны с репортерным геном Потоколы трансформации, а также протоколы для введения нуклеотидных последовательностей в растения могут варьировать в зависимости от типа растения или клетки растения, т.е. однодольного или двудольного, намеченного для трансформации. Подходящие способы введения ДНК-конструкции включают микроинъекцию (Crossway et al. (1986) Biotechniques 4:320-334 и патент США Нуклеотидные конструкции могут быть введены в растения путем введения в контакт растения с вирусом или вирусными нуклеиновыми кислотами. В целом, такие способы включают встраивание нуклеотидной конструкции по данному изобретению в молекулу вирусной ДНК или РНК. Кроме того, признано, что пригодные промоторы охватывают промоторы, используемые для транскрипции вирусными РНК полимеразами. Способы введения нуклеотидных конструкций в растения и экспрессирования белка, закодированного в них, включения молекул вирусных ДНК или РНК, известны в данной области. См., например, патенты США В некоторых вариантах осуществления может быть желательна транзиторная экспрессия. В этих случаях можно использовать стандартные методики транзиторной трансформации. Такие способы включают, но без ограничения, способы вирусной трансформации и микроинъекцию ДНК или РНК, а также другие способы, хорошо известные в данной области. Клетки из растений, которые имеют стабильно включенную нуклеотидную последовательность, можно выращивать в растениях согласно традиционным способам. См., например, McCormick et al. (1986) Plant Cell Reports 5:81-84. Затем эти растения можно выращивать и опылять или тем же трансформированным сортом, или другими сортами, и полученный гибрид будет обладать конститутивной экспрессией желательной фенотипической характеристики, обеспеченной нуклеотидной последовательностью интереса, и/или генетическими маркерами, содержащимися в сайте-мишени или кассете переноса. Для обеспечения стабильного поддержания и наследования экспрессии желаемой фенотипической характеристики можно вырастить два или более поколения, а затем собрать семена, чтобы удостовериться, что достигнута экспрессия желаемой фенотипической характеристики. ПРИМЕРЫ ПРИМЕР 1 Выделение генов-предшественников геномных микроРНК Последовательности, кодирующие гены микроРНК маиса, как описано у Zhang B, et al. (2006) FEBS Lett. 580:3753-62, использовали в качестве запросов для BLAST анализа коллекции Pioneer Unicorn 6.0 маркерных экспрессирующихся последовательностей. Приблизительно 30% запросов имели точные соответствия в коллекции Unicorn 6.0. Следующие праймеры (приобретенные у MWG-BIOTECH Inc.) предназначались для амплификации выбора пяти из этих последовательностей (см. Таблица 1). Таблица 1Праймеры для амплификации предшественников геномных микроРНК ПраймерПоследовательность праймераSEQ ID NO 159cs5'-ccatggcttttcatagcacctctatacctc-3' 1159ca 5'-ggatccacgggcgctcgctgcacccagatcc-3' 2164hs 5'-ggatcctgcgaagctgagtgcagacgtccg-3' 3 164ha5'-ccatgggtacgagggacgatgggattaggc-3' 4168as 5'-ggatccggttcgcgcggagggaaggagggag-3' 5168aa 5'-ccatgggccaatcggctacttgatctcttcccc-3' 6169rs 5'-ggatccctccacacagagaagcaaagaaacc-3' 7169ra 5'-ccatgggtaacctatcgtctattcattttg-3' 8396hs 5'-ggatccgtcccccagatttgctaggacacc-3' 9396ha 5'-ccatggtgggcctgctactatgatgtttag-3' 10 159 смысловой праймер (159cs, SEQ ID NO:1) включал нуклеотиды, которые кодировали Nco I сайт. 159 антисмысловой праймер (159ca, SEQ ID NO:2) включал нуклеотиды, которые кодировали Bam HI сайт. Остальные четыре смысловых праймера (SEQ ID NO: 3, 5, 7 и 9) включали нуклеотиды, которые кодировали Bam HI сайт. Остальные четыре антисмысловых праймера (SEQ ID NO: 4, 6, 8 и 10) включали нуклеотиды, которые кодировали Nco I сайт. Семена Zea mays cv. B73 прорастили и геномную ДНК получили из ткани проростков с помощью набора Qiagen DNeasy Plant Maxi согласно инструкциям производителя. ДНК продукты амплифицировали с помощью геномной ДНК в качестве матрицы и вышеуказанных праймерных пар ExTaq полимеразой (TaKaRa Bio Inc.). Полученные ДНК продукты клонировали в pCR2.1 (Invitrogen) и полностью секвенировали. Охарактеризованные предшественники микроРНК приведены в Таблице 2. Таблица 2Последовательности предшественников микроРНКпредшественник микроРНКSEQ ID NO Длина (нуклеотиды)159c 11875 164h12780 168a13 791169r 14859 396h15 633ПРИМЕР 2 Конструкция искусственных микроРНК последовательностей Искусственные микроРНК (имиРНК), которые будут обладать способностью к сайленсингу гена фитоендесатуразы маиса (NCBI номер U37285), сконструировали главным образом согласно правилам, описанным у Schwab R, et al. (2005) Dev Cell 8 : 517-27. В итоге, микроРНК последовательности имеют длину 21 нуклеотид, начинаются с их 5'-конца с ПРИМЕР 3 Конструкция искусственных последовательностей, отмеченных штрихом
ПРИМЕР 4 Конверсия предшественников геномных микроРНК в предшественники искусственных микроРНК Гены предшественников геномных миРНК конвертировали в имиРНК с помощью перекрывающейся ПЦР (полимеразная цепная реакция), а полученные ДНК полностью секвенировали. Затем эти ДНК клонировали в 5'-3' направлении подходящего промотора в вектор, способный к трансформации маиса. Затем полученные плазмиды были совместно интегрированы в штамм Agrobacterium LBA4404 vir плазмидой PHP10523 (PCT публикация Альтернативно, имиРНК могут быть синтезированы коммерчески, например, с помощью Codon Devices (Cambridge, MA). Затем синтезированную ДНК клонируют в 5'-3' направлении подходящего промотора в вектор, способный к трансформации маиса. Затем полученные плазмиды совместно интегрируют в штамм Agrobacterium LBA4404 плазмидой PHP10523 и могут использовать для трансформации маиса. ПРИМЕР 5 Конверсия предшественников геномных микроРНК в предшественники искусственных микроРНК Гены предшественников геномных миРНК конвертировали в предшественники искусственных миРНК с помощью перекрывающейся ПЦР, как описано в примере 4, а полученные ДНК полностью секвенировали. Получили следующие пять предшественников искусственных миРНК (см. таблицу 4): Таблица 4Последовательности предшественников искусственных микроРНК, нацеливающиеся на фитоендесатуразу предшественник микроРНК SEQ ID NOдлина (нуклеотиды) 159c-PDS22 684164h-PDS 23739 168a-PDS24 791169r-PDS 25860 396h-PDS26633Затем эти ДНК клонировали в 5'-3' направлении ubiquitin промотор-интрона PHP23576. PHP23576 включает Gateway (Invitrogen) L1 и L2 сайты. Затем полученные плазмиды рекомбинировали с плазмидой PHP20622 (PCT публикация ПРИМЕР 6 Трансформация маиса A. Опосредованная частицами доставка ДНК в маисе ДНК-конструкцию можно ввести в клетки маиса, способные расти на приемлемой маисовой культуральной среде. Такие компетентные клетки могут быть из маисовой суспензионной культуры, каллюсной культуры на твердой среде, свежевыделенных незрелых зародышей или меристематическими клетками. Незрелые зародыши Hi-II генотипа можно использовать как клетки-мишени. Початки собирают через приблизительно 10 дней после опыления, 1,2-1,5 мм незрелые зародыши выделяют из зерен и помещают щитком зародыша вниз на маисовую культуральную среду. Незрелые зародыши бомбардируют в течение 18-72 часов после сбора из початка. Между 6 и 18 часами до бомбардировки незрелые зародыши помещают на среду с дополнительным осмотически активным веществом (основная среда MS, Musashige и Skoog, 1962, Physiol. Plant 15:473-497, с 0,25 M сорбитола). Зародыши на высокоосмотической среде используют как мишень для бомбардировки и оставляют на этой среде еще на 18 часов после бомбардировки. Для бомбардировки частицами плазмидную ДНК (описанную выше) осаждают на 1,8 мм вольфрамовые частицы с помощью стандартной CaCl2-спермидиновой химии (см., например, Klein et al., 1987, Nature 327:70-73). Каждый планшет бомбардируют один раз при 600 ПСИ с помощью DuPont Helium Gun (Lowe et al., 1995, Bio/Technol 13:677-682). Для типичных составов среды, используемых для выделения незрелых зародышей маиса, каллюсной инициации, каллюсной пролиферации и регенерации растений, см. Armstrong, C., 1994, в В течение 1-7 дней после бомбардировки частицами зародыши перемещают на культуральную среду на основе среды N6, включающую 3 мг/л селективного агента биалофоса. Зародыши и поздний каллюс переносят на свежие планшеты для отбора каждые 2 недели. Каллюс, развивающийся из незрелых зародышей, обследуют на желаемый фенотип. Через 6-8 недель трансформированный каллюс восстанавливают. B. Трансформация маиса с использованием Agrobacterium Опосредованную Agrobacterium трансформацию маиса выполняют в основном как описано Zhao et al., в Meth. Mol. Biol. 318:315-323 (2006) (см. также Zhao et al., Mol. Breed. 8:323-333 (2001) и патент США 1. Приготовление незрелых зародышей: Незрелые зародыши маиса иссекают из зерновок и помещают в 2 мл микропробирку, включающую 2 мл PHI-A среды. 2. Инфекция Agrobacterium и совместная культивация незрелых зародышей: 2.1 Этап инфекции: PHI-A среду (1) удаляют 1 мл микропипеткой и добавляют 1 мл суспензии Agrobacterium. Пробирку осторожно переворачивают для смешивания. Смесь инкубируют в течение 5 минут при комнатной температуре. 2.2 Этап совместного культивирования: Суспензию Agrobacterium удаляют из этапа инфекции 1 мл микропипеткой. С помощью стерильного шпателя зародыши выскребают из пробирки и переносят на пластину PHI-B среды в 100×15 мм чашке Петри. Зародыши ориентируют осью зародыша вниз на поверхности среды. Пластины с зародышами культивировали при 20°C в темноте в течение трех дней. В фазе совместной культивации можно использовать L-цистеин. Со стандартным бинарным вектором среда для совместного культивирования, дополненная 100-400 мг/л L-цистеина, является критической для восстановления стабильных трансгенных событий. 3. Отбор предполагаемых трансгенных событий: На каждую пластину PHI-D среды в 100×15 мм чашке Петри переносят 10 зародышей, поддерживая ориентацию, и чашки закупоривают парафином. Пластины инкубируют в темноте при 28°C. Активно растущие предполагаемые события, такие как бледно-желтая зародышевая ткань, как ожидается, будут видны через шесть-восемь недель. Зародыши, которые не дают событий, могут быть коричневыми и некротичными, и выражен небольшой рост рыхлой ткани. Предполагаемую трансгенную зародышевую ткань пересевают на свежие PHI-D пластины при двух-трехнедельных интервалах в зависимости от скорости роста. События регистрируют. 4. Регенерация T0 растений: Зародышевую ткань, размноженную на PHI-D среде, пересевают на PHI-E среду (среда созревание соматических зародышей) в 100×25 мм чашки Петри и инкубируют при 28°C в темноте до созревания соматических зародышей, в течение от примерно десяти до восемнадцати дней. Отдельные зрелые соматические зародыши с хорошо определенным щитком и колеоптилем переносят на PHI-F среду проращивания зародышей и инкубируют при 28°C на свету (примерно 80 мкE от холодно-белых или эквивалентных флуоресцентных ламп). На седьмой-десятый день регенерированные растения, высотой примерно 10 см, высаживают в горшок в садоводческую смесь и закаливают с использованием стандартных садоводческих способов. Среда для трансформации растений: 1. PHI-A: 4 г/л CHU основных солей, 1,0 мл/л 1000X витаминной смеси Эриксона, 0,5 мг/л тиамина HCl, 1,5 мг/л 2,4-D, 0,69 г/л L-пролина, 68,5 г/л сахарозы, 36 г/л глюкозы, pH 5,2. Добавить 100 мкМ ацетосирингона (стерилизованного фильтрацией). 2. PHI-B: PHI-A без глюкозы, увеличить 2,4-D до 2 мг/л, снизить сахарозу до 30 г/л и дополненная 0,85 мг/л нитрата серебра (стерилизованного фильтрацией), 3,0 г/л Gelrite®, 100 мкМ ацетосирингона (стерилизованного фильтрацией), pH 5,8. 3. PHI-C: PHI-B без Gelrite® и ацетосирингона, снизить 2,4-D до 1,5 мг/л и дополненная 8,0 г/л агара, 0,5 г/л буфера 2-[N-морфолино]этан-сульфоновой кислоты (MES), 100 мг/л карбенициллина (стерилизованного фильтрацией). 4. PHI-D: PHI-C дополненная 3 мг/л биалофоса (стерилизованного фильтрацией). 5. PHI-E: 4,3 г/л солей Мурасиге-Скуга (MS), (Gibco, BRL 11117-074), 0,5 мг/л никотиновой кислоты, 0,1 мг/л тиамина HCl, 0,5 мг/л пиридоксина HCl, 2,0 мг/л глицина, 0,1 г/л мио-инозитола, 0,5 мг/л зеатина (Sigma, 6. PHI-F: PHI-E без зеатина, IAA, ABA; снизить сахарозу до 40 г/л; заменяя агар на 1,5 г/л Gelrite®; pH 5,6. Растения можно регенерировать из трансгенного каллюса посредством сначала переноса кластеров ткани на среду N6, дополненную 0,2 мг на литр 2,4-D. Через две недели ткань можно перенести на регенерирующую среду (Fromm et al., Bio/Technology 8:833-839 (1990)). Можно регенерировать трансгенные T0 растения и определить их фенотип. Можно собрать T1 семя. Кроме того, рекомбинантная ДНК-конструкция, содержащая проверенный ген Arabidopsis, может быть введена в инбредную линию маиса или прямой трансформацией, или интрогрессией из отдельно трансформированной линии. Трансгенные растения, или инбредные, или гибридные, можно подвергнуть более интенсивным полевым экспериментам по изучению эффектов экспрессии. ПРИМЕР 7 Испытание PDS фенотипа и результаты Через девять-десять дней после опыления зародыши инфицировали Agrobacterium, как описано выше, и провели отбор на Basta. Осуществили пять различных инфицирований, каждое с помощью Agrobacterium, содержащей одну из пяти имиРНК, описанных в примере пять. Соматические зародыши были регенерированы и сгруппированы в сетку 3×3. Один слой пластин, содержащих сгруппированные проростки, подвергли ~114 мЭйнштейн м-2 сек.-1 света. Проросток в середине сетки визуально оценили по фенотипу как любой из: 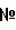 имиРНК% белых растений% светлых % сайленсингаPHP30223 159c -PDS0,014,3 14PHP30448 164h -PDS0,0 7,68 PHP30225168a -PDS нет данныхнет данных нет данныхPHP30451 169r -PDS10,9 47,358 PHP30452396h -PDS 13,059,376 PHP33006396h -PDS 248,3%41,4% 90PHP33007 396h -PDS 318,3% 68,4%87 имиРНК% белых растений% светлых % сайленсингаPHP30223 159c -PDS0,014,3 14PHP30448 164h -PDS0,0 7,68 PHP30225168a -PDS нет данныхнет данных нет данныхPHP30451 169r -PDS10,9 47,358 PHP30452396h -PDS 13,059,376 PHP33006396h -PDS 248,3%41,4% 90PHP33007 396h -PDS 318,3% 68,4%87 Эти результаты показывают, что некоторые из предшественников имиРНК способны продуцировать имиРНК, которые эффективны в генном сайленсинге. Две дополнительных миРНК, нацеленные на фитоендесатуразу, клонировали в 396h остов вместе с их последовательностями, отмеченными штрихом. Обе конструкции показали эффективность сайленсинга, подобную PHP30452, показанной в Таблице 5. ПРИМЕР 8 Нозерн-блот анализ РНК получили из замороженной ткани проростка с помощью Trizol (Invitrogen) согласно протоколу, предоставленному производителем. Общую РНК прогнали на 15% TBE(трисборатный буфер)-мочевина геле PAGE (анализ методом электрофореза в полиакриламидном геле), прогон при 200 В в 0,5xTBE в течение приблизительно 60-90 минут, а затем перенесли на положительно заряженную нейлоновую мембрану BrightStar-Plus (Ambion) с помощью Trans-Blot SD камеры. После переноса блот промыли с 0,5XTBE, и РНК была поперечно связана с мембраной с помощью одного цикла на Auto-Energy в Stratalinker (Stratagene). Блот предварительно гибридизовали в гибридизационном буфере ULTRAhyb-oligo (Ambion) в течение 30 минут при 42°C, а затем гибридизовали в течение ночи при 42°С в гибридизационном буфере ULTRAhyb-oligo с добавлением меченного биотином зонда. Меченный биотином зонд был конкатемером двух копий обратного комплемента последовательности имиРНК, отделенной спейсером длиной 4 пар оснований, и был помечен с помощью набора псорален-биотин неизотопного мечения BrightStar (Ambion). После гибридизации блот промыли раствором для промывания NorthernMax (Ambion), а определение выполнили с помощью набора неизотопного определения BrightStar BioDetect (Ambion) согласно протоколу, предоставленному производителем. Во всех случаях присутствие последовательности имиРНК длиной 21 пар оснований коррелировало с фенотипом. Кроме того, умеренные уровни сигнала коррелировали с бледными растениями, в то время как более высокие уровни сигнала коррелировали с белыми растениями. ПРИМЕР 9 Усеченные предшественники искусственных микроРНК способны к генному сайленсингу В попытке определения способности конструкций, которые содержат предшественники искусственных микроРНК менее чем полной длины, к генному сайленсингу, с помощью ПЦР получили следующие укороченные предшественники имиРНК (см. таблицы 6, 7). Таблица 6Усеченные предшественники искусственных микроРНК, нацеленные на фитоендесатуразу предшественник микроРНК SEQ ID NOдлина (нуклеотиды) 169r-PDS-sht27 96169r-PDS-med 28305 396h-PDS-sht29 117396h-PDS-med 30292Эти укороченные имиРНК-конструкции полностью секвенировали и клонировали в конструкции, как описано в Примере 5. Эти конструкции трансформировали в зародыши маиса и оценили по их способности к сайленсингу PDS. Таблица 7Эффективность сайленсинга усеченных имиРНК Конструкция имиРНК% белых растений% светлых % сайленсингаPHP32347 169r-PDS-sht0,00 0,00%0 PHP32346169r- PDS-med 0,0033,933,9 PHP32349396h- PDS-sht0,000,00 0PHP32348 396h- PDS-med5,2 29,334,5 Эти результаты показывают, что укороченные конструкции способны продуцировать имиРНК. Однако если конструкция слишком укорочена, он уже не способен продуцировать имиРНК. ПРИМЕР 10 Искусственные микроРНК для того, чтобы заставить молчать fad2-1 Вышеприведенные примеры показывают сайленсинг PDS гена маиса, но специалисту в данной области известно, что имиРНК могут быть построены для того, чтобы заставить многие гены. В качестве примера другого гена, которого можно заставить молчать, построили искусственную миРНК, нацеленную на fad2-1, как описано в Примере 2, искусственные последовательности, отмеченные штрихом, построили, как описано в Примере 3, а предшественники имиРНК были созданы, как пояснялось в Примере 4. Эти конструкции приведены в таблице 8. Таблица 8Предшественники искусственных микроРНК, нацеленные на жирнокислотную десатуразу предшественник микроРНК SEQ ID NOдлина (нуклеотиды) 159c-FAD31 684168a-FAD 32790 169r-FAD33 861396h-FAD 34633Затем эти ДНК клонировали в 5'-3' направлении 16 кДа промотора oleosin гена Zea mays, включающего 5' UTR (нетранслируемый участок), в плазмиде PHP20790. PHP20790 включает Gateway (Invitrogen) L1 и L2 сайты. Полученные плазмиды затем рекомбинировали с плазмидой PHP20622. Полученные плазмиды затем совместно интегрировали в штамм Agrobacterium LBA4404 плазмидой PHP10523 и использовали для трансформации маисовых зародышей из культивара GS3, опыленного пыльцой от культивара HC69. Растениям позволили регенерировать и скрестили с использованием трансгенного растения как женской особи и HC69 как мужской особи. Семена собрали и проанализировали на содержание жирных кислот стандартными способами. Анализ ГХ (газовой хроматографии) FAME (жирнокислотного метилового эфира) использовали для исследования, изменяет ли экспрессия искусственной миРНК жирнокислотный профиль семени маиса. Приблизительно проанализировали 50 семян на событие и проанализировали 18-25 событий на конструкцию. Добавили гексан, 1,5 мл, к измельченному маису в лотке для экстракции. Гексан с растворенным маслом перенесли в 1,8 мл стеклянные сосуды для ГХ, в которые добавили 100 мкл триметилсульфония гидроксида для переэтерификации. Жирнокислотные метиловые сложные эфиры (1 мкл инъецировали из гексанового слоя, индекс разведения 80 к 1) отделили и количественно определили с помощью газового хроматографа Agilent 6890, оснащенного 15 м капиллярной колонкой Zebron ZB-wax, диаметр отверстия 0,25 мм, толщина пленки 0,25 мкм. (Каталожный Конструкции с 168a остовом и 396h остовом давали эффективный сайленсинг. Конструкции, выполненные содержащими 159c остов и 169r остов, не дают никакого сайленсинга (% сайленсинга fad2-1; который является процентом событий, показывающих сайленсинг). Последующие поколения трансгенных семян выращивали в поле, а семена T2 и T3 испытали на содержание жирных кислот. Как было показано, эффект этих конструкций наследственный и стабильный. При сравнении с сайленсингом, достигнутым с помощью имиРНК, нацеленной на PDS (% сайленсинга PDS в Таблице 9), оказалось, что некоторые остовы предшественников миРНК молчат во многих типах тканей (396h и потенциально 164h), тогда как другие являются более специфическими по тканям, в которых они эффективны (159c и 169r). Все из пяти миРНК-остовов показали эффективность, по меньшей мере, в одном типе ткани. Таблица 9Предшественники искусственных миРНК, проявляющие избирательную эффективность ОстовSEQ ID NO% сайленсинга PDS % сайленсинга fad2-1159c 22, 3114 0164h 23, нет8нет данных 168a24, 32 нет данных85 169r25, 33 580 396h26, 3476 92Возможные объяснения результатов, представленных в Таблице 9, включают, но без ограничения, дифференциальную стабильность предшественников миРНК временным, пространственным или специфическим для органов способами; дифференциальный процессинг предшественников миРНК (основание для этапов дифференциального пост-транскрипционного процессинга для миРНК обнаружено в животных системах Obernosterer et al. (2007) RNA 12: 1161-1167); или дифференциальную конкуренцию за dicer комплекс среди популяций миРНК. ПРИМЕР 11 Искусственные микроРНК для того, чтобы заставить молчать летальную пятнистость листьев Вышеприведенные примеры показывают сайленсинг PDS гена маиса и fad2 гена, но специалисту в данной области известно, что имиРНК можно построить для того, чтобы заставить молчать несколько генов. В качестве примера другого гена, которого можно заставить молчать, была построена имиРНК, нацеливающаяся на летальную пятнистость листьев, как описано в Примере 2, микроРНК представляет собой 5'-uuugaaaggacgguguaaggc-3' (ДНК-последовательность, соответствующая этой имиРНК, предствлена в SEQ ID NO:35). Искусственные последовательности, отмеченные штрихом, построены, как описано в Примере 3, и показаны в Таблице 10. Предшественники имиРНК создали, как пояснено в Примере 4. Таблица 10Последовательности, отмеченные штрихом, искусственных микроРНК предшественник имиРНК последовательность, отмеченная штрихомSEQ ID NO 168a-llsaccttaatccgtcctttcaaa 3669r-lls cccttacacctgtcctttcaac 37396h-lls gccttacagcgtcctttcaaa38Растения маиса, в которых заставили молчать ген летальной пятнистости листьев (NCBI number, U77345; Gray J et al. (1997) A novel suppressor of cell death in plants encoded by the Lls1 gene of maize. Cell. Apr 4; 89(1):25-31.), проявляют фенотип смертельных повреждений во всех развивающихся листьях, а в некоторых случаях смерть растения. % сайленсинга определили визуальным осмотром растений через приблизительно 8 недель после того, как растения были перенесены в теплицу. Все три конструкции дали эффективный сайленсинг (Таблица 11). Таблица 11Конструкции искусственных миРНК эффективно заставляют молчать летальную пятнистость листьевномер PHP конструкция% сайленсинг PHP36155168a-lls 82PHP 33788 69r-lls80 PHP 36154396h-lls 85ПРИМЕР 12 Искусственные микроРНК как для десатуразы жирной кислоты, так и для белка устойчивости к нескольким лекарственным средствам Иногда желательно заставить молчать более чем один ген с заданной конструкцией. Отдельные предшественники имиРНК могут быть функционально связаны с одним и тем же или с разными промоторами. Альтернативно, два или более предшественников имиРНК могут быть функционально связаны друг с другом, а затем связаны с одним промотором. Из такой конструкции могут быть получены две или более имиРНК. Конструкции, включающие искусственные микроРНК для того, чтобы заставить молчать fad2-1, показаны в примере 10. Как пример другого гена, которого можно заставить молчать, построили искусственную миРНК, нацеливающуюся на белок устойчивости к нескольким лекарственным средствам (MRP), как описано в Примере 2, микроРНК представляет собой 5'-uaauucacaaucucaccacuc-3' (ДНК-последовательность, соответствующая этой имиРНК представлена в SEQ ID NO:39). Искусственные последовательности, отмеченные штрихом, построены, как описано в Примере 3, и показаны в Таблице 12. Создали два предшественника MRP имиРНК, 168a-MRP (SEQ ID No:42) и 396h-MRP (SEQ ID No:43), как пояснялось в Примере 4. Сайленсинг MRP приводит к снижению фитиновой кислоты (Shi J. et al. (2007) Embryo-specific silencing of a transporter reduces phytic acid content of maize and soybean seeds. Nat Biotechnol. Aug;25(8):930-7). Таблица 12Последовательности, отмеченные штрихом,искусственных микроРНК предшественник имиРНК последовательность, отмеченная штрихом SEQ ID NO168a-MRP aagtggctagattgtgaatta40 396h-MRPgagtggtgacattgtgaatta 41Эти предшественники клонировали или в 3'-5' направлении, или в 5'-3' направлении предшественников fad 2 (описанных в Примере 10) для создания кассет, и затем клонировали в конструкции и перенесли в Agrobacterium для создания конструкций, описанных в Таблице 13, как описано ранее. Трансформацию маиса выполнили, как описано в Примере 6, и трансгенные семена будут оценены на содержание фитата и жирных кислот. Таблица 13Конструкции искусственных миРНК, включающие имиРНК, построенные для того, чтобы заставить молчать как fad 2-1, так и MRP номер PHP конструкция PHP38380168aFAD2-1/168a-MRP PHP38381396h-Fad2-1/168a-MRP PHP38382 168a-MRP/396h-FAD2-1PHP38383 396h-MRP/396h-FAD2-1Формула изобретения1. Выделенный фрагмент нуклеиновой кислоты для понижения экспрессии последовательности-мишени, по существу являющейся дезоксирибонуклеотидной последовательностью, представленной в SEQ ID NO: 11, где (i) нуклеотиды 430-450 в SEQ ID NO: 11 замещены первой вариабельной нуклеотидной субпоследовательностью, размер которой варьируется от 19 до 24 нуклеотидов в зависимости от последовательности-мишени, экспрессия которой подлежит понижению, (ii) нуклеотиды 244-264 в SEQ ID NO: 11 замещены второй вариабельной нуклеотидной субпоследовательностью, размер которой варьируется от 19 до 24 нуклеотидов, причем указанная вторая вариабельная нуклеотидная субпоследовательность способна гибридизоваться с первой вариабельной субпоследовательностью предшественника миРНК, и (iii) предшественник миРНК, полученный указанным выделенным фрагментом нуклеиновой кислоты, имеет ту же стеблевую структуру, что и предшественник миРНК, полученный эндогенной последовательностью SEQ ID NO: 11. 2. Рекомбинантная конструкция для понижения экспрессии последовательности-мишени, включающая выделенный фрагмент нуклеиновой кислоты по п.1, функционально связанный, по меньшей мере, с одной регуляторной последовательностью. 3. Клетка маиса для понижения экспрессии последовательности-мишени, включающая рекомбинантную конструкцию по п.2. 4. Способ отбора клеток, имеющих пониженный уровень экспрессии включающий: (а) трансформацию, по меньшей мере, одной растительной клетки маиса рекомбинантной конструкцией по п.2; и(b) отбор той трансформированной растительной клетки (клеток) маиса, у которых уровень экспрессии той последовательности-мишени понижен по сравнению с уровнем экспрессии гена-мишени в растительной клетке маиса дикого типа. Популярные патенты: 2239968 Способ предпосевной обработки семян овощных культур ... обработки семян /Н.А.Болотов, Е.Е.Кашкин, В.Е.Шевченко (RU). - Заявка № 99121038/13; заявлено 05.10.1999; опубл. 20.07.2001, БИ № 20 //Изобретения и полезные модели. - 2001, - № 20).К недостаткам данного способа относятся то, что растворы католит и аналита лишь подавляют развитие на поверхности семян патогенных микроорганизмов и не способствуют энергии роста зародышей.Известно также средство для предпосевной обработки посеян масличных культур, представляющее собой водный раствор действующего начала, в котором в качестве действующего начала используют природный минерал бишофит, а его концентрация в водном растворе составляет 10-15 об.% (RU, патент № 2145471 С1, МПК7 А 01 С 1/00. ... 2403705 Способ автоматического управления температурно-световым режимом в теплице ... алгоритму компьютерный задатчик осуществляет переход на управление температурно-световым режимом по формулам (6) или (7), которые содержат параметр текущего значения вышедшего из-под контроля САУ фактора температурно-светового режима. Сам фактор фиксируется в неизменном состоянии в данном промежутке времени. Использование данного способа автоматического управления температурно-световым режимом в теплице значительно повышает эффективность использования световой энергии солнца и облучательной установки культивируемыми растениями, а значит, позволяет сократить длительность периода вегетации до начала плодоношения, что важно для культивации светокультуры, увеличить ... 2087614 Способ создания травяного газонного покрытия открытых спортивных площадок и ухода за ним ... известный способ не содержит всего комплекса необходимых мер, позволяющего создать качественное травяное покрытие, выдерживающее большие нагрузки при использовании спортивной площадки. Наиболее близким к изобретению является способ создания травяного газонного покрытия, включающий засеивание площадки, полив дождеванием, ручную прополку и регулярное скашивание травы [2] Но рекомендации, описанные в указанном способе также не достаточны для создания качественного травяного покрытия. Задачей изобретения является улучшение качества травяного газонного покрытия открытых спортивных площадок, сохранение и улучшение его свойств в течение длительного времени, что, в сою очередь, снижает ... 2404581 Способ изготовления муляжей анатомических препаратов полых и трубчатых органов ... были изготовлены 12 коррозионных препаратов желчных протоков с использованием "Протакрила М". Печень погружалась в 35-38%-ную соляную кислоту на 4-5 суток [Панфилов С.А., Фомичев О.М., Мамедли З.З. Трехмерная холангиография при заболеваниях желчных путей. - Россия, М., Лаборатория компьютерных технологий кафедры общей хирургии МГМСУ]. К недостаткам данного полимера можно также отнести быструю полимеризацию, что неудобно при изготовлении органокомплексов больших размеров, а также выраженную ломкость получаемых слепков, высокую стоимость и труднодоступность "Протакрила М". Кроме того, он быстро застывает на воздухе, что не совсем удобно при изготовлении объемных ... 2084132 Устройство для выращивания растений ... общая схема устройства для выращивания растений; фиг. 2 растильня с держателем; фиг. 3 - держатели с горшками; фиг. 4 схема электрической связи горшка (датчиков) с растворным узлом и шкафом управления; фиг. 5 общий вид растильни с системой перемещения горшков в держателе. Устройство для выращивания растений содержит растильню 1, состоящую из держателя 2, имеющего отверстия многоугольной формы 3 (гнезда), в которые вставляют горшки 4 конусообразной формы из эластичного материала с субстратом 5 и рассадой с корневой системой 6. В нижней части растильни 1 в питательные каналы 7 установлены вкладыши 8, 9, 10 со сливными патрубками 11, 12, 13. Под днищем растильни 1 для ... |
Еще из этого раздела: 2204241 Способ определения поливных норм при капельном орошении томатов 2106082 Устройство для укладки подстилочного навоза в бурт 2196418 Устройство для укладки, сушки и хранения прессованного сена и соломы в рулонах 2094986 Гербицидный состав 2182420 Устройство для перерезания стволов деревьев 2434381 Технологическая линия для приготовления и раздачи влажных кормов 2188534 Способ уборки льна-долгунца 2163758 Способ и устройство контроля количества меда в улье 2215407 Способ создания исходного материала для селекции растений 2197082 Установка для охлаждения молока с использованием естественного холода |
Изобретения в сельском хозяйстве
Обработка почвы в сельском и лесном хозяйствах
Посадка, посев, удобрение
Уборка урожая, жатва
Обработка и хранение продуктов полеводства и садоводства
Садоводство, разведение овощей, цветов, риса, фруктов, винограда, лесное хозяйство
Новые виды растений или способы их выращивания
Производство молочных продуктов
Животноводство, разведение и содержание птицы, рыбы, насекомых, рыбоводство, рыболовство
Поимка, отлов или отпугивание животных
Консервирование туш животных, или растений или их частей
Биоцидная, репеллентная, аттрактантная или регулирующая рост растений активность химических соединений или препаратов
Хлебопекарные печи, машины и прочее оборудование для хлебопечения
Машины или оборудование для приготовления или обработки теста
Обработка муки или теста для выпечки, способы выпечки, мучные изделия
|
|
||
 последовательностям, отмеченным штрихом
последовательностям, отмеченным штрихом МикроРНК или миРНК
МикроРНК или миРНК -глюкуронидазы и введены в небобовое Nicotiana tabacum и бобовое Lotus corniculatus, и в обоих случаях активность специфического для корня промотора была сохранена. Leach и Aoyagi (1991) описывают их анализ промоторов высоко экспрессируемых rolC и rolD индуцируемых корнем генов Agrobacterium rhizogenes (см. Plant Science (Limerick) 79(1):69-76). Они заключили, что энхансер и предпочтительные для ткани ДНК детерминанты диссоциированы в этих промоторах. Teeri et al. (1989) использовали генное слияние с lacZ, чтобы показать, что T-ДНК ген Agrobacterium, кодирующий октопинсинтазу, особенно активен в эпидермисе кончика корня, и что TR2' ген специфичен для корня в здоровом растении и стимулируется ранением в листовой ткани, особенно желательна комбинация характеристик для применения с инсектицидным или ларвицидным геном (см. EMBO J. 8(2):343-350). TR1' ген, слитый с nptII (неомицин фосфотрансфераза II), показал подобные характеристики. Дополнительные предпочтительные для корня промоторы включают промотор VfENOD-GRP3 гена (Kuster et al. (1995) Plant Mol. Biol. 29(4):759-772); и rolB промотор (Capana et al. (1994) Plant Mol. Biol. 25(4):681-691. См. также патенты США
-глюкуронидазы и введены в небобовое Nicotiana tabacum и бобовое Lotus corniculatus, и в обоих случаях активность специфического для корня промотора была сохранена. Leach и Aoyagi (1991) описывают их анализ промоторов высоко экспрессируемых rolC и rolD индуцируемых корнем генов Agrobacterium rhizogenes (см. Plant Science (Limerick) 79(1):69-76). Они заключили, что энхансер и предпочтительные для ткани ДНК детерминанты диссоциированы в этих промоторах. Teeri et al. (1989) использовали генное слияние с lacZ, чтобы показать, что T-ДНК ген Agrobacterium, кодирующий октопинсинтазу, особенно активен в эпидермисе кончика корня, и что TR2' ген специфичен для корня в здоровом растении и стимулируется ранением в листовой ткани, особенно желательна комбинация характеристик для применения с инсектицидным или ларвицидным геном (см. EMBO J. 8(2):343-350). TR1' ген, слитый с nptII (неомицин фосфотрансфераза II), показал подобные характеристики. Дополнительные предпочтительные для корня промоторы включают промотор VfENOD-GRP3 гена (Kuster et al. (1995) Plant Mol. Biol. 29(4):759-772); и rolB промотор (Capana et al. (1994) Plant Mol. Biol. 25(4):681-691. См. также патенты США